I
–Я–†–Ш–У–Ю–Ґ–Ю–Т–Ы–Х–Э–Ш–Х –•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–• –Я–†–Х–Я–Р–†–Р–Ґ–Ю–Т
–Ь–Х–Ґ–Ю–Ф –Ф–Р–Т–Ы–Х–Э–Ђ–• –Я–†–Х–Я–Р–†–Р–Ґ–Ю–Т
–Ь–µ—В–Њ–і –і–∞–≤–ї–µ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –≤ –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –Є–Љ–µ–µ—В –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –Є—Б—В–Њ—А–Є—З–µ—Б–Ї–Є–є –Є–љ—В–µ—А–µ—Б –Є –њ—А–Є–Љ–µ–љ—П–µ—В—Б—П —А–µ–і–Ї–Њ, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –Њ–љ —В–µ—Е–љ–Є—З–µ—Б–Ї–Є —Б–ї–Њ–ґ–љ–µ–µ –Є —З–∞—Й–µ –њ—А–Є–≤–Њ–і–Є—В –Ї –∞—А—В–µ—Д–∞–Ї—В–∞–Љ. –І–∞—Б—В—М –Љ–µ—В–∞—Д–∞–Ј–љ—Л—Е –њ–ї–∞—Б—В–Є–љ–Њ–Ї –љ–µ–Љ–Є–љ—Г–µ–Љ–Њ —А–∞–Ј—А—Г—И–∞–µ—В—Б—П, –≤ —В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї –Љ–љ–Њ–≥–Є–µ –Ї–ї–µ—В–Ї–Є –њ–ї–Њ—Е–Њ —А–∞—Б–њ–ї–∞—Б—В—Л–≤–∞—О—В—Б—П, —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л –ї–µ–ґ–∞—В —Б–Ї—Г—З–µ–љ–љ–Њ –Є –љ–µ –≤ –Њ–і–љ–Њ–є –њ–ї–Њ—Б–Ї–Њ—Б—В–Є. –Я—А–Є—Е–Њ–і–Є—В—Б—П –≥–Њ—В–Њ–≤–Є—В—М –±–Њ–ї—М—И–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –њ—А–µ–њ–∞—А–∞—В–Њ–≤, —З—В–Њ–±—Л –Њ—В—Л—Б–Ї–∞—В—М –љ–∞ –љ–Є—Е –Ї–ї–µ—В–Ї–Є, –њ—А–Є–≥–Њ–і–љ—Л–µ –і–ї—П –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П. –≠—В–Є–Љ –Љ–µ—В–Њ–і–Њ–Љ –і–Њ —Б–Є—Е –њ–Њ—А –њ–Њ–ї—М–Ј—Г—О—В—Б—П –ї–Є—И—М –љ–µ–Љ–љ–Њ–≥–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–Є, –Њ–і–љ–∞–Ї–Њ –љ–µ–Ї–Њ—В–Њ—А—Л–µ, –њ—А–Є–±–µ–≥–∞—П –Ї —А–∞–Ј–ї–Є—З–љ—Л–Љ —Г—Б–Њ–≤–µ—А—И–µ–љ—Б—В–≤–Њ–≤–∞–љ–Є—П–Љ, –і–Њ–±–Є–≤–∞—О—В—Б—П —Е–Њ—А–Њ—И–Є—Е —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤. –Т –ї–Є—В–µ—А–∞—В—Г—А–µ –Љ–Њ–ґ–љ–Њ –љ–∞–є—В–Є –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Љ–µ—В–Њ–і–Є–Ї –њ–Њ–ї—Г—З–µ–љ–Є—П –і–∞–≤–ї–µ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –Ш—Е —А–∞–Ј–ї–Є—З–Є—П —Б–≤–Њ–і—П—В—Б—П –Ї –њ—А–Є–Љ–µ–љ–µ–љ–Є—О —А–∞–Ј–љ—Л—Е –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Є—Е —А–∞—Б—В–≤–Њ—А–Њ–≤ (–і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–∞—П –≤–Њ–і–∞, —А–∞—Б—В–≤–Њ—А —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П) –Є–ї–Є —Д–Є–Ї—Б–∞—В–Њ—А–Њ–≤. –Э–Є–ґ–µ –Љ—Л –њ—А–Є–≤–Њ–і–Є–Љ –Љ–µ—В–Њ–і–Є–Ї—Г, –њ–Њ –Ї–Њ—В–Њ—А–Њ–є —А–∞–±–Њ—В–∞–ї–Є –≤ 60-—Е –≥–Њ–і–∞—Е –≤ –ї–∞–±–Њ—А–∞—В–Њ—А–Є–Є –†. –Ь–∞—В—В–µ—П.
–Я—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ –Ї–Њ–ї—Е–Є—Ж–Є–љ–Є—А–Њ–≤–∞–љ–љ–Њ–µ –ґ–Є–≤–Њ—В–љ–Њ–µ –Ј–∞–±–Є–≤–∞—О—В –Є –±–µ—А—Г—В –Ї—Г—Б–Њ—З–Ї–Є —Б–µ–ї–µ–Ј–µ–љ–Ї–Є –Є–ї–Є –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ —А–∞–Ј–Љ–µ—А–Њ–Љ –Њ–Ї–Њ–ї–Њ 1 –Љ–Љ. –Ъ—Г—Б–Њ—З–Ї–Є —В–Ї–∞–љ–Є –њ–Њ–Љ–µ—Й–∞—О—В –љ–∞ 10 –Љ–Є–љ. –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ—Г—О –≤–Њ–і—Г (–≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ), –∞ –Ј–∞—В–µ–Љ –њ–µ—А–µ–љ–Њ—Б—П—В –љ–∞ 20-30 –Љ–Є–љ. –≤ 50%-–љ—Г—О —Г–Ї—Б—Г—Б–љ—Г—О –Ї–Є—Б–ї–Њ—В—Г. –Ф–∞–ї–µ–µ –Ї—Г—Б–Њ—З–µ–Ї —В–Ї–∞–љ–Є –њ–µ—А–µ–љ–Њ—Б—П—В –љ–∞ —З–Є—Б—В–Њ–µ –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ, –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ —Б–Љ–∞–Ј–∞–љ–љ–Њ–µ –Ї—Г—А–Є–љ—Л–Љ –±–µ–ї–Ї–Њ–Љ, –Ј–∞–Ї—А—Л–≤–∞—О—В –њ–Њ–Ї—А–Њ–≤–љ—Л–Љ —Б—В–µ–Ї–ї–Њ–Љ –Є –Є–Ј–Њ –≤—Б–µ—Е —Б–Є–ї –љ–∞–і–∞–≤–ї–Є–≤–∞—О—В –љ–∞ –њ–Њ–Ї—А–Њ–≤–љ–Њ–µ —Б—В–µ–Ї–ї–Њ –±–Њ–ї—М—И–Є–Љ –њ–∞–ї—М—Ж–µ–Љ. –Э–µ–Ї–Њ—В–Њ—А—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–Є –Ј–∞–≤–Њ—А–∞—З–Є–≤–∞—О—В –њ—А–µ–њ–∞—А–∞—В, —В. –µ. –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ —Б –Ї—Г—Б–Њ—З–Ї–Њ–Љ —В–Ї–∞–љ–Є, –Ј–∞–Ї—А—Л—В—Л–Љ –њ–Њ–Ї—А–Њ–≤–љ—Л–Љ —Б—В–µ–Ї–ї–Њ–Љ, –≤ —Д–Є–ї—М—В—А–Њ–≤–∞–ї—М–љ—Г—О –±—Г–Љ–∞–≥—Г –Є –њ–Њ–Љ–µ—Й–∞—О—В –њ–Њ–і –њ—А–µ—Б—Б. –Я–Њ—Б—В–Њ—П–љ–љ–Њ–µ –і–∞–≤–ї–µ–љ–Є–µ –љ–∞ –њ—А–µ–њ–∞—А–∞—В, —Б–Њ–Ј–і–∞–≤–∞–µ–Љ–Њ–µ –≥—А—Г–Ј–Њ–Љ, –Љ–Њ–ґ–µ—В –±—Л—В—М —Г—Б–њ–µ—И–љ–Њ —Г—Б–Є–ї–µ–љ–Њ —Б –њ–Њ–Љ–Њ—Й—М—О —А—Л—З–∞–≥–∞ –і–Њ 75 –Ї–≥.
–Я—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ –њ–µ—А–µ–љ–Њ—Б—П—В –≤ –Ї—О–≤–µ—В—Г —Б 70%-–љ—Л–Љ —Б–њ–Є—А—В–Њ–Љ, –Ј–∞–Ї—А—Л–≤–∞—О—В –Є –Њ—Б—В–∞–≤–ї—П—О—В —В–∞–Љ –љ–∞ 2-3 –і–љ—П. –Т —Б–њ–Є—А—В–µ –њ–Њ–Ї—А–Њ–≤–љ–Њ–µ —Б—В–µ–Ї–ї–Њ –Њ—В–њ–∞–і–∞–µ—В –Є –Њ—Б—В–∞—С—В—Б—П —В–Њ–ї—М–Ї–Њ —В–Ї–∞–љ—М, –њ—А–Є–Ї–ї–µ–µ–љ–љ–∞—П –Ї –њ—А–µ–і–Љ–µ—В–љ–Њ–Љ—Г —Б—В–µ–Ї–ї—Г. –Я–Њ—Б–ї–µ —Н—В–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В –њ—А–Њ–Љ—Л–≤–∞—О—В –і–≤–∞ —А–∞–Ј–∞ –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ. –Т—Б–ї–µ–і –Ј–∞ —Н—В–Є–Љ –њ—А–µ–њ–∞—А–∞—В –≤ —В–µ—З–µ–љ–Є–µ 12 –Љ–Є–љ. –њ–Њ–і–≤–µ—А–≥–∞—О—В –≥–Є–і—А–Њ–ї–Є–Ј—Г –≤ 1%-–љ–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л –≤ —В–µ—А–Љ–Њ—Б—В–∞—В–µ –њ—А–Є 58¬∞–°. –Я–Њ—Б–ї–µ –≥–Є–і—А–Њ–ї–Є–Ј–∞ –њ—А–µ–њ–∞—А–∞—В –і–≤–∞ —А–∞–Ј–∞ –њ—А–Њ–Љ—Л–≤–∞—О—В –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ. –Ю–Ї—А–∞—И–Є–≤–∞—О—В —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л –њ–Њ –Љ–µ—В–Њ–і—Г –§–µ–ї—М–≥–µ–љ–∞ –Є–ї–Є –≥–µ–Љ–∞—В–Њ–Ї—Б–Є–ї–Є–љ–Њ–Љ –њ–Њ –≠—А–ї–Є—Е—Г.
–Ь–Х–Ґ–Ю–Ф –Т–Ђ–°–£–®–Х–Э–Э–Ђ–• –Я–†–Х–Я–Р–†–Р–Ґ–Ю–Т
–° —В–µ—Е –њ–Њ—А, –Ї–∞–Ї –≤ –Ї–Њ–љ—Ж–µ 50-—Е –≥–Њ–і–Њ–≤ –њ–Њ—П–≤–Є–ї–Є—Б—М –њ–µ—А–≤—Л–µ –Њ–њ–Є—Б–∞–љ–Є—П —Н—В–Њ–≥–Њ –Љ–µ—В–Њ–і–∞, –±—Л–ї–Њ –њ—А–µ–і–ї–Њ–ґ–µ–љ–Њ –Њ—З–µ–љ—М –Љ–љ–Њ–≥–Њ –µ–≥–Њ –Љ–Њ–і–Є—Д–Є–Ї–∞—Ж–Є–є. –Т –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –њ–Њ—З—В–Є –Ї–∞–ґ–і—Л–є –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М, –і–ї–Є—В–µ–ї—М–љ–Њ–µ –≤—А–µ–Љ—П —А–∞–±–Њ—В–∞–≤—И–Є–є —Б —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞–Љ–Є, –њ–Њ–ї—М–Ј—Г–µ—В—Б—П —Б–Њ–±—Б—В–≤–µ–љ–љ–Њ–є –≤–∞—А–Є–∞—Ж–Є–µ–є —Н—В–Њ–≥–Њ –Љ–µ—В–Њ–і–∞ –Є–ї–Є –њ—А–Њ–±—Г–µ—В –µ–≥–Њ —А–∞–Ј–ї–Є—З–љ—Л–µ –≤–Є–і–Њ–Є–Ј–Љ–µ–љ–µ–љ–Є—П. –Э–Є–ґ–µ –њ—А–Є–≤–Њ–і–Є—В—Б—П –Љ–µ—В–Њ–і–Є–Ї–∞, –њ–Њ –Ї–Њ—В–Њ—А–Њ–є –Љ—Л —А–∞–±–Њ—В–∞–ї–Є –њ–Њ—Б–ї–µ–і–љ–µ–µ –≤—А–µ–Љ—П. –°–ї–µ–і—Г–µ—В –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –Њ–њ–Є—Б–∞–љ–љ–∞—П –Љ–µ—В–Њ–і–Є–Ї–∞, —В–Њ—З–љ–µ–µ —В–∞ –µ—С —З–∞—Б—В—М, –Ї–Њ—В–Њ—А–∞—П –Ї–∞—Б–∞–µ—В—Б—П –Ї–Њ–ї—Е–Є—Ж–Є–љ–Є—А–Њ–≤–∞–љ–Є—П in vivo, –Љ–Њ–ґ–µ—В –±—Л—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–∞ –њ—А–Є —А–∞–±–Њ—В–µ —В–Њ–ї—М–Ї–Њ —Б –Љ–µ–ї–Ї–Є–Љ–Є –Ј–≤–µ—А—М–Ї–∞–Љ–Є –Є–ї–Є –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–Љ–Є —Б—А–µ–і–љ–Є—Е —А–∞–Ј–Љ–µ—А–Њ–≤ (–љ–∞–њ—А–Є–Љ–µ—А, –і–Њ —Б–Њ–±–Њ–ї—П –њ–ї–Є –±–Њ–±—А–∞ –≤–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ). –Я—А–Є —Н—В–Њ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М –і–Њ–ї–ґ–µ–љ —А–∞—Б–њ–Њ–ї–∞–≥–∞—В—М –ґ–Є–≤—Л–Љ–Є –Ј–≤–µ—А—П–Љ–Є, –Ї–Њ—В–Њ—А—Л–Љ –≤–≤–Њ–і—П—В –≤–љ—Г—В—А–Є–±—А—О—И–Є–љ–љ–Њ –Ї–Њ–ї—Е–Є—Ж–Є–љ –Є –Ї–Њ—В–Њ—А—Л—Е —З–µ—А–µ–Ј –љ–µ–Ї–Њ—В–Њ—А–Њ–µ –≤—А–µ–Љ—П –Ј–∞–±–Є–≤–∞—О—В. –Т —В–µ—Е —Б–ї—Г—З–∞—П—Е, –Ї–Њ–≥–і–∞ –њ—А–µ–і—Б—В–Њ–Є—В –Є—Б—Б–ї–µ–і–Њ–≤–∞—В—М —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–є –љ–∞–±–Њ—А –Ј–≤–µ—А–µ–є –Ї—А—Г–њ–љ—Л—Е —А–∞–Ј–Љ–µ—А–Њ–≤ –Є–ї–Є –Є–Љ–µ–µ—В—Б—П —Б–≤–µ–ґ–Є–є —В—А—Г–њ–љ—Л–є –Љ–∞—В–µ—А–Є–∞–ї, –њ–Њ–ї—М–Ј—Г—О—В—Б—П –Є–љ—Л–Љ–Є —Б–њ–Њ—Б–Њ–±–∞–Љ–Є –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ (–Ї—А–∞—В–Ї–Њ–≤—А–µ–Љ–µ–љ–љ–∞—П –Ї—Г–ї—М—В—Г—А–∞ –Ї–ї–µ—В–Њ–Ї –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞, –Ї—Г–ї—М—В—Г—А–∞ –ї–µ–є–Ї–Њ—Ж–Є—В–Њ–≤ –Ї—А–Њ–≤–Є –Є –і—А.). –Ю–і–љ–∞–Ї–Њ –≤–Њ –≤—Б–µ—Е —Б–ї—Г—З–∞—П—Е –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ–µ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –љ–∞—З–Є–љ–∞—П —Б –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –љ–∞ –Ї–ї–µ—В–Ї–Є, –Њ—Б—В–∞—С—В—Б—П –≤ —Б—Г—Й–љ–Њ—Б—В–Є –љ–µ–Є–Ј–Љ–µ–љ–љ—Л–Љ. –Ъ—А–Њ–≤–µ—В–≤–Њ—А–љ—Л–µ –Њ—А–≥–∞–љ—Л вАУ –Ї–Њ—Б—В–љ—Л–є –Љ–Њ–Ј–≥ –Є —Б–µ–ї–µ–Ј—С–љ–Ї–∞ —Г–і–Њ–±–љ—Л –≤ —В–Њ–Љ –Њ—В–љ–Њ—И–µ–љ–Є–Є, —З—В–Њ –Љ–µ–ґ–Ї–ї–µ—В–Њ—З–љ—Л–µ —Б–≤—П–Ј–Є –≤ –љ–Є—Е –Њ—З–µ–љ—М —Б–ї–∞–±—Л–µ. –Т –і—А—Г–≥–Є—Е —Б–ї—Г—З–∞—П—Е (–љ–∞–њ—А–Є–Љ–µ—А, –њ—А–Є–≥–Њ—В–Њ–≤–ї—П—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –Є–Ј —А–Њ–≥–Њ–≤–Є—Ж—Л –≥–ї–∞–Ј–∞) —В–Ї–∞–љ—М –њ—А–Є—Е–Њ–і–Є—В—Б—П –Є–Ј–Љ–µ–ї—М—З–∞—В—М –Љ–µ—Е–∞–љ–Є—З–µ—Б–Ї–Є (–Є –њ—А–Є —Н—В–Њ–Љ –Љ–љ–Њ–≥–Є–µ –Ї–ї–µ—В–Ї–Є —А–∞–Ј—А—Г—И–∞—О—В—Б—П) –Є–ї–Є —А–∞–Ј—А—Л–≤–∞—В—М –Љ–µ–ґ–Ї–ї–µ—В–Њ—З–љ—Л–µ —Б–≤—П–Ј–Є —Д–µ—А–Љ–µ–љ—В–∞–Љ–Є (—В—А–Є–њ—Б–Є–љ–Њ–Љ). –Э–Є–ґ–µ –Њ–њ–Є—Б–∞–љ–∞ –Љ–µ—В–Њ–і–Є–Ї–∞ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Ј –Ї–ї–µ—В–Њ–Ї –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞. –° –љ–µ–Ј–љ–∞—З–Є—В–µ–ї—М–љ—Л–Љ–Є –Є–Ј–Љ–µ–љ–µ–љ–Є—П–Љ–Є –Њ–љ–∞ –њ—А–Є–≥–Њ–і–љ–∞ –і–ї—П –ї—О–±–Њ–є –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Є –∞–Ї—В–Є–≤–љ–Њ–є —В–Ї–∞–љ–Є.
1. –Ц–Є–≤–Њ—В–љ–Њ–Љ—Г –≤–љ—Г—В—А–Є–±—А—О—И–Є–љ–љ–Њ –≤–≤–µ—Б—В–Є 0,04%-–љ—Л–є —А–∞—Б—В–≤–Њ—А –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞ –≤ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ 0,01 –Љ–ї –љ–∞ 1 –≥ –≤–µ—Б–∞.
2. –І–µ—А–µ–Ј 1 —З–∞—Б –ґ–Є–≤–Њ—В–љ–Њ–µ –Ј–∞–±–Є—В—М, –±—Л—Б—В—А–Њ –Є–Ј–≤–ї–µ—З—М –±–µ–і—А–µ–љ–љ—Л–µ –Ї–Њ—Б—В–Є, –Њ—З–Є—Б—В–Є—В—М –Є—Е –Њ—В –њ—А–Є–ї–µ–≥–∞—О—Й–Є—Е –Љ—Л—И—Ж –Є –Њ—В—А–µ–Ј–∞—В—М —Н–њ–Є—Д–Є–Ј—Л. –Ъ–Њ—Б—В–љ—Л–є –Љ–Њ–Ј–≥ –Є–Ј –±–µ–і—А–µ–љ–љ–Њ–є –Ї–Њ—Б—В–Є –≤—Л–Љ—Л—В—М —В—С–њ–ї—Л–Љ (37¬∞–°) –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Є–Љ —А–∞—Б—В–≤–Њ—А–Њ–Љ –≤ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Г—О –њ—А–Њ–±–Є—А–Ї—Г, –Є—Б–њ–Њ–ї—М–Ј—Г—П –і–ї—П —Н—В–Њ–є —Ж–µ–ї–Є –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Є–є —И–њ—А–Є—Ж —Б –Є–≥–ї–Њ–є. –Т –Ї–∞—З–µ—Б—В–≤–µ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –Љ–Њ–ґ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М 0,9%-–љ—Л–є —А–∞—Б—В–≤–Њ—А —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П (—В—А–µ—Е–Ј–∞–Љ–µ—Й—С–љ–љ—Л–є –ї–Є–Љ–Њ–љ–љ–Њ–Ї–Є—Б–ї—Л–є –љ–∞—В—А–Є–є) –Є–ї–Є 0,56%-–љ—Л–є —А–∞—Б—В–≤–Њ—А —Е–ї–Њ—А–Є—Б—В–Њ–≥–Њ –Ї–∞–ї–Є—П.
3. –Ъ—Г—Б–Њ—З–Ї–Є –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –Є–Ј–Љ–µ–ї—М—З–Є—В—М —Б –њ–Њ–Љ–Њ—Й—М—О –њ–∞—Б—В–µ—А–Њ–≤—Б–Ї–Њ–є –њ–Є–њ–µ—В–Ї–Є –Є–ї–Є –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ —И–њ—А–Є—Ж–∞ —Б –Є–≥–ї–Њ–є, –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —А–∞–Ј —Н–љ–µ—А–≥–Є—З–љ–Њ –≤—Б–∞—Б—Л–≤–∞—П –Є –≤—Л–і—Г–≤–∞—П –ґ–Є–і–Ї–Њ—Б—В—М. –Я–Њ–ї—Г—З–µ–љ–љ—Г—О –Ї–ї–µ—В–Њ—З–љ—Г—О —Б—Г—Б–њ–µ–љ–Ј–Є—О –Є–љ–Ї—Г–±–Є—А–Њ–≤–∞—В—М –≤ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–є —Б—А–µ–і–µ: –≤ —Б–ї—Г—З–∞–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П вАУ 15-30 –Љ–Є–љ. –Є –≤ –≤–∞—А–Є–∞–љ—В–µ —Б –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ —Е–ї–Њ—А–Є–і–∞ –Ї–∞–ї–Є—П вАУ 5 –Љ–Є–љ. –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞–°.
4. –Ю—В—Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞—В—М –њ–Њ–ї—Г—З–µ–љ–љ—Г—О —Б—Г—Б–њ–µ–љ–Ј–Є—О –≤ —В–µ—З–µ–љ–Є–µ 5 –Љ–Є–љ. –њ—А–Є 800-1000 –Њ–±/–Љ–Є–љ. –Ю—Б–∞–і–Ї–∞ –≤ –њ—А–Њ–±–Є—А–Ї–µ –і–Њ–ї–ґ–љ–Њ –±—Л—В—М –љ–µ–Љ–љ–Њ–≥–Њ (—В–Њ–љ–Ї–Є–є —Б–ї–Њ–є –љ–∞ –і–љ–µ –њ—А–Њ–±–Є—А–Ї–Є).
5. –Ю—Б—В–Њ—А–Њ–ґ–љ–Њ —Б–ї–Є—В—М –Є–ї–Є –Њ—В—Б–Њ—Б–∞—В—М –њ–Є–њ–µ—В–Ї–Њ–є –≤—Б—О –љ–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М, —Б—В–∞—А–∞—П—Б—М –њ—А–Є —Н—В–Њ–Љ –љ–µ –њ–Њ—В—А–µ–≤–Њ–ґ–Є—В—М –Њ—Б–∞–і–Ї–∞. –Ъ–ї–µ—В–Ї–Є —Д–Є–Ї—Б–Є—А–Њ–≤–∞—В—М —Б–Љ–µ—Б—М—О –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ —Б –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є (3 —З–∞—Б—В–Є –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є 1 —З–∞—Б—В—М —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л). –Т–Љ–µ—Б—В–Њ –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Љ–Њ–ґ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М —Н—В–Є–ї–Њ–≤—Л–є —Б–њ–Є—А—В. –Ъ –Њ—Б–∞–і–Ї—Г –Њ—Б—В–Њ—А–Њ–ґ–љ–Њ –њ—А–Є–ї–Є—В—М —Б–≤–µ–ґ–µ–њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–є –Є –Њ—Е–ї–∞–ґ–і–µ–љ–љ—Л–є —Д–Є–Ї—Б–∞—В–Њ—А. –§–Є–Ї—Б–∞—В–Њ—А–∞ –љ–∞–ї–Є–≤–∞—О—В –Њ–Ї–Њ–ї–Њ 2 –Љ–ї, —Б—В–∞—А–∞—П—Б—М –љ–µ —А–∞–Ј–±–Є—В—М –Њ—Б–∞–і–Њ–Ї. –Ю—З–µ–љ—М —З–∞—Б—В–Њ —Д–Є–Ї—Б–∞—Ж–Є—О –Њ—Б–∞–і–Ї–∞ –љ–∞—З–Є–љ–∞—О—В –Є–љ–∞—З–µ. –§–Є–Ї—Б–∞—В–Њ—А –њ—А–Є–ї–Є–≤–∞—О—В –Њ—З–µ–љ—М –Љ–µ–і–ї–µ–љ–љ–Њ, –њ–Њ –Њ–і–љ–Њ–є –Ї–∞–њ–ї–µ, –Є —Н–љ–µ—А–≥–Є—З–љ–Њ –≤—Б—В—А—П—Е–Є–≤–∞—О—В –њ—А–Њ–±–Є—А–Ї—Г, —З—В–Њ–±—Л –і–Њ–±–Є—В—М—Б—П –Њ–і–љ–Њ—А–Њ–і–љ–Њ–є –≤–Ј–≤–µ—Б–Є –Ї–ї–µ—В–Њ–Ї –≤ —Д–Є–Ї—Б–∞—В–Њ—А–µ. –Ь—Л –Є—Б–њ—А–Њ–±–Њ–≤–∞–ї–Є –Є —Б—А–∞–≤–љ–Є–≤–∞–ї–Є –Њ–±–∞ —Б–њ–Њ—Б–Њ–±–∞ —Д–Є–Ї—Б–∞—Ж–Є–Є, –њ–Њ –Ј–∞—В—А—Г–і–љ—П–µ–Љ—Б—П –Њ—В–і–∞—В—М –Ї–∞–Ї–Њ–Љ—Г-–ї–Є–±–Њ –Є–Ј –љ–Є—Е –њ—А–µ–і–њ–Њ—З—В–µ–љ–Є–µ. –Т –њ–Њ–ї–µ–≤—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е —Г–і–Њ–±–µ–љ –њ–µ—А–≤—Л–є —Б–њ–Њ—Б–Њ–±, —В–∞–Ї –Ї–∞–Ї –њ—А–Є –љ–µ–Љ —Б–Њ–Ї—А–∞—Й–∞–µ—В—Б—П —З–Є—Б–ї–Њ –њ–Њ—Б–ї–µ–і—Г—О—Й–Є—Е —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–є.
6. –Ю–±—Й–µ–µ –≤—А–µ–Љ—П —Д–Є–Ї—Б–∞—Ж–Є–Є 1-2 —З–∞—Б–∞. –Ю–і–љ–∞–Ї–Њ —Д–Є–Ї—Б–Є—А–Њ–≤–∞—В—М –Є –і–∞–ґ–µ —Е—А–∞–љ–Є—В—М –≤–Ј–≤–µ—Б—М –Ї–ї–µ—В–Њ–Ї –≤ —Д–Є–Ї—Б–∞—В–Њ—А–µ –љ–∞ —Е–Њ–ї–Њ–і—Г –Љ–Њ–ґ–љ–Њ –і–Њ-–≤–Њ–ї—М–љ–Њ –і–Њ–ї–≥–Њ, –µ—Б–ї–Є –Є—Б–Ї–ї—О—З–Є—В—М –≤—Л—Б—Л—Е–∞–љ–Є–µ —Д–Є–Ї—Б–∞—В–Њ—А–∞. –Ч–∞ –≤—А–µ–Љ—П —Д–Є–Ї—Б–∞—Ж–Є–Є –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ —Б–Љ–µ–љ–Є—В—М 2-3 —А–∞–Ј–∞ —Д–Є–Ї—Б–∞—В–Њ—А, —Б –њ—А–Њ–Љ–µ–ґ—Г—В–Њ—З–љ—Л–Љ —А–µ—Б—Г—Б–њ–µ–љ–Ј–Є—А–Њ–≤–∞–љ–Є–µ–Љ –Њ—Б–∞–і–Ї–∞ –Є –њ–Њ—Б–ї–µ–і—Г—О—Й–Є–Љ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ–Љ. –Х—Б–ї–Є –Њ—Б–∞–і–Њ–Ї —Б —Б–∞–Љ–Њ–≥–Њ –љ–∞—З–∞–ї–∞ —Д–Є–Ї—Б–Є—А–Њ–≤–∞–ї—Б—П —Ж–µ–ї–Њ–є –≥–ї—Л–±–Ї–Њ–є, —В–Њ —Б—В–∞—А—Л–є —Д–Є–Ї—Б–∞—В–Њ—А –Љ–Њ–ґ–љ–Њ –Њ—Б—В–Њ—А–Њ–ґ–љ–Њ –Њ—В—Б–Њ—Б–∞—В—М –њ–Є–њ–µ—В–Ї–Њ–є –Є –љ–∞–ї–Є—В—М —Б–≤–µ–ґ–Є–є –±–µ–Ј —А–µ—Б—Г—Б–њ–µ–њ–Ј–Є—А–Њ–≤–∞–љ–Є—П.
7. –Т –њ–Њ—Б–ї–µ–і–љ–µ–Љ —Д–Є–Ї—Б–∞—В–Њ—А–µ –Њ—Б–∞–і–Њ–Ї —А–µ—Б—Г—Б–њ–µ–љ–і–Є—А—Г–µ—В—Б—П. –§–Є–Ї—Б–∞—В–Њ—А–∞ –љ–∞–ї–Є—В—М –љ–µ–Љ–љ–Њ–≥–Њ (–Њ–±—Л—З–љ–Њ 0,5 –Љ–ї) —В–∞–Ї, —З—В–Њ–±—Л –њ–Њ–ї—Г—З–Є–ї–∞—Б—М –Љ–Њ–ї–Њ—З–љ–Њ–≥–Њ —Ж–≤–µ—В–∞ —Б–ї–µ–≥–Ї–∞ –Њ–њ–∞–ї–µ—Б—Ж–Є—А—Г—О—Й–∞—П –≤–Ј–≤–µ—Б—М.
8. –Э–∞ —З–Є—Б—В–Њ –≤—Л–Љ—Л—В–Њ–µ –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ (—Б—В–µ–Ї–ї–∞ –і–µ—А–ґ–∞—В—М –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї–µ) –љ–∞–љ–µ—Б—В–Є 2-3 –Ї–∞–њ–ї–Є —Б—Г—Б–њ–µ–љ–Ј–Є–Є –Ї–ї–µ—В–Њ–Ї –≤ —Д–Є–Ї—Б–∞—В–Њ—А–µ. –°—В–µ–Ї–ї–Њ –±—Л—Б—В—А–Њ –њ—А–Њ–≤–µ—Б—В–Є —З–µ—А–µ–Ј –њ–ї–∞–Љ—П –≥–Њ—А–µ–ї–Ї–Є, —З—В–Њ–±—Л —Д–Є–Ї—Б–∞—В–Њ—А –≤–Њ—Б–њ–ї–∞–Љ–µ–љ–Є–ї—Б—П, –љ–Њ –љ–µ –і–Њ–њ—Г—Б–Ї–∞—В—М –њ–µ—А–µ–≥—А–µ–≤–∞–љ–Є—П. –§–Є–Ї—Б–∞—В–Њ—А –њ—А–Є —Н—В–Њ–Љ –≤—Л–≥–Њ—А–∞–µ—В, –∞ –Ї–ї–µ—В–Ї–Є –њ—А–Њ—З–љ–Њ —Д–Є–Ї—Б–Є—А—Г—О—В—Б—П –Ї —Б—В–µ–Ї–ї—Г. –Ы–Є—И–љ—П—П –≤–ї–∞–≥–∞ –≤—Л—Б—Г—И–Є–≤–∞–µ—В—Б—П —Д–µ–љ–Њ–Љ –Є–ї–Є —Б–і—Г–≤–∞–µ—В—Б—П –≥—А—Г—И–µ–є.
9. –•–Њ—А–Њ—И–Њ –≤—Л—Б—Г—И–µ–љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –Њ–Ї—А–∞—И–Є–≤–∞—О—В—Б—П —А–∞–Ј–ї–Є—З–љ—Л–Љ–Є –Ї—А–∞—Б–Є—В–µ–ї—П–Љ–Є, –≤ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В —Ж–µ–ї–Є, —Б—В–Њ—П—Й–µ–є –њ–µ—А–µ–і –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–µ–Љ (—Б–Љ. —А–∞–Ј–і–µ–ї ¬Ђ–†–∞–Ј–ї–Є—З–љ—Л–µ —Б–њ–Њ—Б–Њ–±—Л –Њ–Ї—А–∞—Б–Ї–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤¬ї).
–Ъ–∞—З–µ—Б—В–≤–Њ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Ј–∞–≤–Є—Б–Є—В –Њ—В —В—Й–∞—В–µ–ї—М–љ–Њ—Б—В–Є –њ–Њ–і–≥–Њ—В–Њ–≤–Ї–Є –њ—А–µ–і–Љ–µ—В–љ—Л—Е —Б—В—С–Ї–Њ–ї. –Я—А–µ–і–Љ–µ—В–љ—Л–µ —Б—В–µ–Ї–ї–∞ –≤—Л–Љ—Л—В—М –≤–∞—В–љ—Л–Љ —В–∞–Љ–њ–Њ–љ–Њ–Љ –≤ –Љ—Л–ї—М–љ–Њ–є –≤–Њ–і–µ –Є –њ–Њ—Б—В–∞–≤–Є—В—М –њ—А–Њ–Ї–Є–њ—П—В–Є—В—М –≤ –≤–Њ–і–µ —Б –і–µ—В—Б–Ї–Є–Љ –Љ—Л–ї–Њ–Љ –Є–ї–Є —Б—В–Є—А–∞–ї—М–љ—Л–Љ –њ–Њ—А–Њ—И–Ї–Њ–Љ 10-15 –Љ–Є–љ. –Ъ–∞–ґ–і–Њ–µ —Б—В–µ–Ї–ї–Њ –Њ—В–і–µ–ї—М–љ–Њ –њ—А–Њ–Љ—Л—В—М –≤ –љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е —Б–Љ–µ–љ–∞—Е –≥–Њ—А—П—З–µ–є –≤–Њ–і—Л –і–Њ –њ–Њ–ї–љ–Њ–≥–Њ –Њ—В–Љ—Л–≤–∞–љ–Є—П –Љ—Л–ї–∞. –•–Њ—А–Њ—И–Њ –Њ—В–Љ—Л—В—Л–µ —Б—В–µ–Ї–ї–∞ –њ–Њ–Љ–µ—Б—В–Є—В—М –≤ —А–∞—Б—В–≤–Њ—А —Е—А–Њ–Љ–њ–Є–Ї–∞ –љ–∞ 8-12 —З–∞—Б. –Ч–∞ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —З–∞—Б–Њ–≤ –і–Њ —Д–Є–Ї—Б–∞—Ж–Є–Є —Б—В–µ–Ї–ї–∞ –Њ—В–Љ—Л—В—М –Њ—В —Е—А–Њ–Љ–њ–Є–Ї–∞ –≤ –≤–Њ–і–Њ–њ—А–Њ–≤–Њ–і–љ–Њ–є –≤–Њ–і–µ –Є –≤ –љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е —Б–Љ–µ–љ–∞—Е –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л –Є –≤ –њ–Њ—Б–ї–µ–і–љ–µ–є –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ —Б—В–µ–Ї–ї–∞ –њ–Њ—Б—В–∞–≤–Є—В—М –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї.
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–Х –Я–†–Х–Я–Р–†–Р–Ґ–Ђ –Ш–Ч –Ъ–†–Р–Ґ–Ъ–Ю–Т–†–Х–Ь–Х–Э–Э–Ю–Щ –Ъ–£–Ы–ђ–Ґ–£–†–Ђ –Ъ–Ы–Х–Ґ–Ю–Ъ
–Ъ—А–∞—В–Ї–Њ–≤—А–µ–Љ–µ–љ–љ–∞—П –Ї—Г–ї—М—В—Г—А–∞ –Ї–ї–µ—В–Њ–Ї –Є –Ї–Њ–ї—Е–Є—Ж–Є–љ–Є—А–Њ–≤–∞–љ–Є–µ in vitro –і–∞—О—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –њ—А–Є–≥–Њ—В–Њ–≤–Є—В—М —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –≤ —В–∞–Ї–Є—Е —Б–ї—Г—З–∞—П—Е, –Ї–Њ–≥–і–∞ –Њ–њ–Є—Б–∞–љ–љ–∞—П –≤—Л—И–µ –Љ–µ—В–Њ–і–Є–Ї–∞ –љ–µ–њ—А–Є–Љ–µ–љ–Є–Љ–∞. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ —Г–і–∞—С—В—Б—П –њ–Њ–ї—Г—З–Є—В—М —Е–Њ—А–Њ—И–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –Є–Ј –њ—Г–љ–Ї—В–∞—В–∞ –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞, –≤–Ј—П—В–Њ–≥–Њ –њ—А–Є–ґ–Є–Ј–љ–µ–љ–љ–Њ, –њ—А–Є –Њ—В—Б—В—А–µ–ї–∞—Е –ґ–Є–≤–Њ—В–љ—Л—Е, –∞ —В–∞–Ї–ґ–µ –≤–Њ–Њ–±—Й–µ –Є–Ј –ї—О–±—Л—Е –Ј–∞–±–Є—В—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е, –њ–Њ—Б–ї–µ —Б–Љ–µ—А—В–Є –Ї–Њ—В–Њ—А—Л—Е –њ—А–Њ—И–ї–Њ –Њ—В 1 —З–∞—Б–∞ –і–Њ –і–µ—Б—П—В–Ї–Њ–≤ —З–∞—Б–Њ–≤ (–≤ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В —В–µ–Љ–њ–µ—А–∞—В—Г—А—Л, –њ—А–Є –Ї–Њ—В–Њ—А–Њ–є —Е—А–∞–љ–Є–ї—Б—П —В—А—Г–њ). –Ь–∞–ї–∞—П —В—А—Г–і–Њ—С–Љ–Ї–Њ—Б—В—М –Є –±—Л—Б—В—А–Њ—В–∞ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –∞ —В–∞–Ї–ґ–µ –љ–µ–љ—Г–ґ–љ–Њ—Б—В—М —Б–Њ–±–ї—О–і–µ–љ–Є—П —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є –њ–Њ–Ј–≤–Њ–ї—П—О—В –њ—А–Є–Љ–µ–љ—П—В—М —Н—В—Г –Љ–µ—В–Њ–і–Є–Ї—Г –≤ –њ–Њ–ї–µ–≤—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е.
–Ы—Г—З—И–Є–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –і–∞—С—В –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞, –Є–Ј–≤–ї–µ—З—С–љ–љ–Њ–≥–Њ –Є–Ј –≥—А—Г–і–Є–љ—Л –њ—А–Є –њ–Њ–Љ–Њ—Й–Є –Ї–Њ—Б—В–љ–Њ-–Љ–Њ–Ј–≥–Њ–≤–Њ–є –Є–≥–ї—Л. –Я–Њ–ї—Г—З–µ–љ–Є—О —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Ј –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –±–µ–і—А–µ–љ–љ—Л—Е –Ї–Њ—Б—В–µ–є —Г –Ї—А—Г–њ–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е —З–∞—Б—В–Њ –Љ–µ—И–∞–µ—В –љ–∞–ї–Є—З–Є–µ –ґ–Є—А–∞.
–Я—Г–љ–Ї—В–∞—В –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ (0,5-1 –Љ–ї) –њ–Њ–Љ–µ—Й–∞—О—В –≤ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Л–µ –њ—А–Њ–±–Є—А–Ї–Є, —Б–Њ–і–µ—А–ґ–∞—Й–Є–µ 9 –Љ–ї –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і—Л 199, 1 –Љ–ї –≥–Њ—В–Њ–≤–Њ–є —Б—Л–≤–Њ—А–Њ—В–Ї–Є –Ї—А—Г–њ–љ–Њ–≥–Њ —А–Њ–≥–∞—В–Њ–≥–Њ —Б–Ї–Њ—В–∞, –Є –Є–љ–Ї—Г–±–Є—А—Г—О—В –Њ—В 1 –і–Њ 5 —З–∞—Б., –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞. –Ф–ї—П —Н—В–Њ–є —Ж–µ–ї–Є –Љ–Њ–ґ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М —В–µ—А–Љ–Њ—Б—В–∞—В, –≤–Њ–і—П–љ—Г—О –±–∞–љ—О –Є–ї–Є –њ—А–Њ—Б—В–Њ —В–µ—А–Љ–Њ—Б —Б —В—С–њ–ї–Њ–є –≤–Њ–і–Њ–є. –Ч–∞ 40 –Љ–Є–љ. –і–Њ –Ї–Њ–љ—Ж–∞ –Є–љ–Ї—Г–±–∞—Ж–Є–Є –≤–≤–Њ–і—П—В 0,2 –Љ–ї 0,004%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞. –Я–Њ –Њ–Ї–Њ–љ—З–∞–љ–Є–Є –Є–љ–Ї—Г–±–∞—Ж–Є–Є –Ї–ї–µ—В–Ї–Є –Њ—Б–∞–ґ–і–∞—О—В —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ–Љ –≤ —В–µ—З–µ–љ–Є–µ 5 –Љ–Є–љ. –њ—А–Є 800 –Њ–±/–Љ–Є–љ. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Б–ї–Є–≤–∞—О—В, –∞ –Њ—Б–∞–і–Њ–Ї —А–∞–Ј–±–Є–≤–∞–µ—В—Б—П —Б–Є–ї—М–љ–Њ–є —Б—В—А—Г—С–є –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ (0,56%-–љ—Л–є —Е–ї–Њ—А–Є—Б—В—Л–є –Ї–∞–ї–Є–є), –≤—Л–і—Г–≤–∞–µ–Љ–Њ–≥–Њ –Є–Ј —И–њ—А–Є—Ж–∞ –Є–ї–Є –њ–Є–њ–µ—В–Ї–Є. –У–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–∞—П –Њ–±—А–∞–±–Њ—В–Ї–∞ –Є –≤—Б–µ –і–∞–ї—М–љ–µ–є—И–Є–µ –њ—А–Њ—Ж–µ–і—Г—А—Л –њ—А–Њ–≤–Њ–і—П—В—Б—П —В–∞–Ї –ґ–µ, –Ї–∞–Ї –Є –≤ —Б–ї—Г—З–∞–µ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Ј –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –ґ–Є–≤–Њ—В–љ—Л—Е, –Ї–Њ—В–Њ—А—Л–Љ –і–Њ –Ј–∞–±–Њ—П –≤–≤–Њ–і–Є–ї–Є –Ї–Њ–ї—Е–Є—Ж–Є–љ (—Б–Љ. –≤—Л—И–µ).
–•—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –Є–Ј —Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ –Љ–∞—В–µ—А–Є–∞–ї–∞, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –њ–Њ–ї—Г—З–∞—О—В –њ—А–Є –≤–≤–µ–і–µ–љ–Є–Є –µ–≥–Њ –≤ –Ї—Г–ї—М—В—Г—А—Г. –Э–Њ –µ—Б–ї–Є –љ–µ—В —Г—Б–ї–Њ–≤–Є–є –і–ї—П –Ї—Г–ї—М—В—Г—А—Л —В–Ї–∞–љ–µ–є, —В–Њ –Љ–Њ–ґ–љ–Њ –њ–Њ–ї—Г—З–Є—В—М —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –Є–Ј —Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ–Њ–є —В–Ї–∞–љ–Є.
–Р–Ї—В–Є–≤–љ–Њ –і–µ–ї—П—Й–Є–µ—Б—П —В–Ї–∞–љ–Є —Н–Љ–±—А–Є–Њ–љ–Њ–≤ (–њ–µ—З–µ–љ—М, —Б–µ–ї–µ–Ј—С–љ–Ї–∞ –Є –і—А.) –Є–ї–Є —Ж–µ–ї—Л–є —Н–Љ–±—А–Є–Њ–љ –љ–∞ —А–∞–љ–љ–Є—Е —Б—В–∞–і–Є—П—Е —А–∞–Ј–≤–Є—В–Є—П –њ–Њ–Љ–µ—Й–∞—О—В –≤ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Г—О –њ—А–Њ–±–Є—А–Ї—Г, —Б–Њ–і–µ—А–ґ–∞—Й—Г—О –њ–Є—В–∞—В–µ–ї—М–љ—Г—О —Б—А–µ–і—Г –Ш–≥–ї–∞ —Б –≥–ї—Г—В–∞–Љ–Є–љ–Њ–Љ (–Є–ї–Є —Б—А–µ–і—Г 199), –Є —А–µ—Б—Г—Б–њ–µ–љ–Ј–Є—А—Г—О—В –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Є–Љ —И–њ—А–Є—Ж–µ–Љ —Б —В–Њ–ї—Б—В–Њ–є –Є–≥–ї–Њ–є. –Ъ–ї–µ—В–Ї–Є –Њ—Б—В–∞—О—В—Б—П –≤ —Б—Г—Б–њ–µ–љ–Ј–Є–Є –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞–° –≤ —В–µ—З–µ–љ–Є–µ 1-1,5 —З–∞—Б. –Ґ–∞–Ї–ґ–µ –Ј–∞ 40 –Љ–Є–љ. –і–Њ –Ї–Њ–љ—Ж–∞ –Є–љ–Ї—Г–±–Є—А–Њ–≤–∞–љ–Є—П –≤–≤–Њ–і—П—В 0,2 –Љ–ї 0,004 %-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞.
–Ч–∞—В–µ–Љ –Ї–ї–µ—В–Ї–Є –Њ—Б–∞–ґ–і–∞—О—В —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ–Љ –Є –≤—Б—О –і–∞–ї—М–љ–µ–є—И—Г—О –Њ–±—А–∞–±–Њ—В–Ї—Г –њ—А–Њ–≤–Њ–і—П—В —В–∞–Ї –ґ–µ, –Ї–∞–Ї –Є –≤ —Б–ї—Г—З–∞–µ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Ј –Ї–ї–µ—В–Њ–Ї –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞.
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–Х –Я–†–Х–Я–Р–†–Р–Ґ–Ђ –Ш–Ч –Ъ–£–Ы–ђ–Ґ–£–†–Ђ –Ы–Х–Щ–Ъ–Ю–¶–Ш–Ґ–Ю–Т –Я–Х–†–Ш–§–Х–†–Ш–І–Х–°–Ъ–Ю–Щ –Ъ–†–Ю–Т–Ш
–Ъ—А–∞—В–Ї–Њ –Љ–µ—В–Њ–і–Є–Ї–∞ —Б–≤–Њ–і–Є—В—Б—П –Ї —В–Њ–Љ—Г, —З—В–Њ –њ–ї–∞–Ј–Љ—Г –Ї—А–Њ–≤–Є —Б –ї–µ–є–Ї–Њ—Ж–Є—В–∞–Љ–Є —Б–Љ–µ—И–Є–≤–∞—О—В —Б –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і–Њ–є –Є –і–Њ–±–∞–≤–ї—П—О—В —Д–Є—В–Њ–≥–µ–Љ–∞–≥–≥–ї—О—В–Є–љ–Є–љ (–§–У–Р), –±–ї–∞–≥–Њ–і–∞—А—П –Љ–Є—В–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Њ–Љ—Г –і–µ–є—Б—В–≤–Є—О –Ї–Њ—В–Њ—А–Њ–≥–Њ —З–µ—А–µ–Ј 24 —З–∞—Б–∞ –≤ –Ї—Г–ї—М—В—Г—А–µ –њ–Њ—П–≤–ї—П—О—В—Б—П –њ–µ—А–≤—Л–µ –Љ–Є—В–Њ–Ј—Л. –Э–∞–Є–±–Њ–ї—М—И–µ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –і–µ–ї—П—Й–Є—Е—Б—П –Ї–ї–µ—В–Њ–Ї, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –љ–∞–±–ї—О–і–∞–µ—В—Б—П —З–µ—А–µ–Ј 72 —З–∞—Б–∞ –њ–Њ—Б–ї–µ –≤–≤–µ–і–µ–љ–Є—П –≤ –Ї—Г–ї—М—В—Г—А—Г (Moorhead et al., 1960). –Т –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ –≤–Ј—П—В–Њ–є –Ї—А–Њ–≤–Є —А–∞–Ј–ї–Є—З–∞—О—В –Љ–∞–Ї—А–Њ- –Є –Љ–Є–Ї—А–Њ–Љ–µ—В–Њ–і.
–Ь–Р–Ъ–†–Ю–Ь–Х–Ґ–Ю–Ф
1. –Ч–∞–≥–Њ—В–∞–≤–ї–Є–≤–∞—О—В —Б—В–µ—А–Є–ї—М–љ—Л–µ –њ—А–Њ–±–Є—А–Ї–Є, —Б–Њ–і–µ—А–ґ–∞—Й–Є–µ 0,5 –Љ–ї —А–∞–±–Њ—З–µ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –≥–µ–њ–∞—А–Є–љ–∞ (10 –Љ–ї —А–∞—Б—В–≤–Њ—А–∞ –•–µ–љ–Ї—Б–∞ +0,5 –Љ–ї —А–∞—Б—В–≤–Њ—А–∞ –≥–µ–њ–∞—А–Є–љ–∞). –Я—А–Њ–±–Є—А–Ї—Г –Ј–∞–Ї—А—Л–≤–∞—О—В —Б—В–µ—А–Є–ї—М–љ–Њ–є –њ–µ–љ–Є—Ж–Є–ї–ї–Є–љ–Њ–≤–Њ–є –њ—А–Њ–±–Ї–Њ–є. –Х—С –Ї—А–∞—П –Ј–∞–Ї–ї–µ–Є–≤–∞—О—В –ї–µ–є–Ї–Њ–њ–ї–∞—Б—В—Л—А–µ–Љ.
2. –®–њ—А–Є—Ж, –њ—А–Њ–Ї–Є–њ—П—З–µ–љ–љ—Л–є –≤ —В–µ—З–µ–љ–Є–µ 40 –Љ–Є–љ., –њ—А–Њ–Љ—Л—В—М —З–Є—Б—В—Л–Љ –≥–µ–њ–∞—А–Є–љ–Њ–Љ –Є –≤ –љ–µ–≥–Њ –±—А–∞—В—М –Ї—А–Њ–≤—М.
3. –Ъ—А–Њ–≤—М –±–µ—А—Г—В –Є–Ј —П—А–µ–Љ–љ–Њ–є –≤–µ–љ—Л –ґ–Є–≤–Њ—В–љ–Њ–≥–Њ —И–њ—А–Є—Ж–µ–Љ –Є –њ–µ—А–µ–ї–Є–≤–∞—О—В –≤ –њ—А–Њ–±–Є—А–Ї—Г —Б –≥–µ–њ–∞—А–Є–љ–Њ–Љ, –њ—А–Њ–Ї–Њ–ї–Њ–≤ –њ–µ–љ–Є—Ж–Є–ї–ї–Є–љ–Њ–≤—Г—О –њ—А–Њ–±–Ї—Г –Є–≥–ї–Њ–є.
4. –°—В–∞–≤—П—В –њ—А–Њ–±–Є—А–Ї–Є –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї –њ—А–Є +4¬∞–° –љ–∞ 30 –Љ–Є–љ. –Х—Б–ї–Є –Ј–∞ —Н—В–Њ –≤—А–µ–Љ—П —Н—А–Є—В—А–Њ—Ж–Є—В—Л –љ–µ –Њ—Б–µ–ї–Є, —В–Њ –≤ —Б—В–µ—А–Є–ї—М–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –і–Њ–±–∞–≤–ї—П—О—В 10%-–љ—Л–є —А–∞—Б—В–≤–Њ—А –ґ–µ–ї–∞—В–Є–љ—Л –≤ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–Є 3:1 (3 —З–∞—Б—В–Є –Ї—А–Њ–≤–Є: 1 —З–∞—Б—В—М –ґ–µ–ї–∞—В–Є–љ—Л). –≠—В—Г –Є —А—П–і –њ–Њ—Б–ї–µ–і—Г—О—Й–Є—Е –њ—А–Њ—Ж–µ–і—Г—А –њ—А–Њ–≤–Њ–і—П—В –≤ —Б—В–µ—А–Є–ї—М–љ–Њ–Љ –±–Њ–Ї—Б–µ, –Ї–Њ—В–Њ—А—Л–є –±—Л–ї –Њ–±–ї—Г—З—С–љ –±–∞–Ї—В–µ—А–Є—Ж–Є–і–љ—Л–Љ–Є –ї–∞–Љ–њ–∞–Љ–Є –≤ —В–µ—З–µ–љ–Є–µ 1,5 —З–∞—Б. –њ–µ—А–µ–і –њ–Њ—Б—В–∞–љ–Њ–≤–Ї–Њ–є –Ї—Г–ї—М—В—Г—А—Л.
5. –Я–Њ—Б–ї–µ —В–Њ–≥–Њ, –Ї–∞–Ї —Н—А–Є—В—А–Њ—Ж–Є—В—Л –Њ—Б—П–і—Г—В, –њ–ї–∞–Ј–Љ—Г –Њ—В—Б–∞—Б—Л–≤–∞—О—В –Љ–µ—А–љ–Њ–є –њ–Є–њ–µ—В–Ї–Њ–є.
6. –Я–Њ–ї—Г—З–µ–љ–љ—Г—О –њ–ї–∞–Ј–Љ—Г —Б–Љ–µ—И–Є–≤–∞—О—В —Б –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і–Њ–є (—Б—А–µ–і–∞ 199 –Є–ї–Є –Ш–≥–ї–∞ —Б –≥–ї—Г—В–∞–Љ–Є–љ–Њ–Љ) –≤ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–Є 1 —З–∞—Б—В—М –њ–ї–∞–Ј–Љ—Л –Є 3 —З–∞—Б—В–Є –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і—Л.
7. –Э–∞ 10 –Љ–ї —Н—В–Њ–є —Б–Љ–µ—Б–Є –і–Њ–±–∞–≤–ї—П—О—В 0,2 –Љ–ї —Д–Є—В–Њ–≥–µ–Љ–∞–≥–≥–ї—О—В–Є–љ–Є–љ–∞-–Ь —Д–Є—А–Љ—Л Wellcome (–Є–ї–Є Difco) –Є–ї–Є 0,02 –Љ–ї —Д–Є—В–Њ–≥–µ–Љ–∞–≥–≥–ї—О—В–Є–љ–Є–љ–∞-–† —Д–Є—А–Љ—Л Difco. –Ф–Њ–±–∞–≤–ї—П—О—В –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Є (–њ–µ–љ–Є—Ж–Є–ї–ї–Є–љ –Є–Ј —А–∞—Б—З—С—В–∞ 100 –µ–і. –љ–∞ 1 –Љ–ї, —Б—В—А–µ–њ—В–Њ–Љ–Є—Ж–Є–љ вАУ 50 –µ–і. –љ–∞ 1 –Љ–ї).
8. –Я–Њ–ї—Г—З–µ–љ–љ—Г—О –≤–Ј–≤–µ—Б—М –Ї–ї–µ—В–Њ–Ї –Љ–Њ–ґ–љ–Њ –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞—В—М –≤ —З–∞—И–Ї–∞—Е –Ъ–Њ—А—А–µ–ї—П, –≤ –Љ–∞—В—А–∞—Б–∞—Е, –≤ –њ–ї–Њ—Б–Ї–Њ–і–Њ–љ–љ—Л—Е –Ї–Њ–ї–±–∞—Е –Є–ї–Є –≤ –њ–µ–љ–Є—Ж–Є–ї–ї–Є–љ–Њ–≤—Л—Е —Д–ї–∞–Ї–Њ–љ–∞—Е (–њ–Њ 1,5 –Љ–ї –љ–∞ —Д–ї–∞–Ї–Њ–љ). –Т–Ј–≤–µ—Б—М –Ї–ї–µ—В–Њ–Ї —А–∞–Ј–ї–Є–≤–∞—О—В –њ–Њ —Д–ї–∞–Ї–Њ–љ–∞–Љ, –Ј–∞–Ї—А—Л–≤–∞—О—В —А–µ–Ј–Є–љ–Њ–≤—Л–Љ–Є –њ—А–Њ–±–Ї–∞–Љ–Є –Є –Њ—Б—В–∞–≤–ї—П—О—В –≤ —В–µ—А–Љ–Њ—Б—В–∞—В–µ –њ—А–Є 37¬∞–° –љ–∞ 72 —З–∞—Б–∞. –І–∞—Б—В—М –њ–ї–∞–Ј–Љ—Л –Љ–Њ–ґ–љ–Њ —Б–Њ—Е—А–∞–љ–Є—В—М –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї–µ –Є –≤–≤–µ—Б—В–Є –≤ –Ї—Г–ї—М—В—Г—А—Г —Б–њ—Г—Б—В—П 3-4 –і–љ—П, –Ї–Њ–≥–і–∞ —Б—В–∞–љ—Г—В –Є–Ј–≤–µ—Б—В–љ—Л —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–µ—А–≤–Њ–є –Ї—Г–ї—М—В—Г—А—Л.
9. –Ч–∞ 1-1,5 —З–∞—Б–∞ –і–Њ —Д–Є–Ї—Б–∞—Ж–Є–Є –≤ –Ї—Г–ї—М—В—Г—А—Г –≤–≤–Њ–і—П—В –њ–Њ–і–Њ–≥—А–µ—В—Л–є –і–Њ 37¬∞–° —А–∞—Б—В–≤–Њ—А –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞, —Б–Њ–і–µ—А–ґ–∞—Й–Є–є 20 –≥–∞–Љ–Љ –љ–∞ 1 –Љ–ї –≤–Њ–і—Л (0,002%-–љ—Л–є —А–∞—Б—В–≤–Њ—А), –Є–Ј —А–∞—Б—З—С—В–∞ 0,5 –≥–∞–Љ–Љ –љ–∞ 1 –Љ–ї —Б—А–µ–і—Л1.( 1 –Т —Н—В–Њ–є –Є –њ–Њ—Б–ї–µ–і—Г—О—Й–Є—Е –њ—А–Њ—Ж–µ–і—Г—А–∞—Е –љ–µ —В—А–µ–±—Г–µ—В—Б—П —Б–Њ–±–ї—О–і–∞—В—М —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В—М.)
10. –Я–Њ—Б–ї–µ 1 —З–∞—Б–∞ –Є–љ–Ї—Г–±–∞—Ж–Є–Є —Б –Ї–Њ–ї—Е–Є—Ж–Є–љ–Њ–Љ –Ї—Г–ї—М—В—Г—А—Г —А–∞–Ј–ї–Є–≤–∞—О—В –њ–Њ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Л–Љ –њ—А–Њ–±–Є—А–Ї–∞–Љ –Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А—Г—О—В 5-8 –Љ–Є–љ. –њ—А–Є 1000 –Њ–±/–Љ–Є–љ (—А–µ–ґ–Є–Љ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є—П –њ—А–Є –≤—Б–µ–є –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–є –Њ–±—А–∞–±–Њ—В–Ї–µ —Б–Њ—Е—А–∞–љ—П–µ—В—Б—П –њ—А–Є–Љ–µ—А–љ–Њ —В–∞–Ї–Є–Љ –ґ–µ).
11. –£–і–∞–ї–Є—В—М –љ–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М, –∞ –Ї –Њ—Б–∞–і–Ї—Г –і–Њ–±–∞–≤–Є—В—М —В—С–њ–ї—Л–є (37¬∞–°) —А–∞—Б—В–≤–Њ—А –•–µ–љ–Ї—Б–∞, –≤–Ј–±–Њ–ї—В–∞—В—М –њ–Њ—Б—В—Г–Ї–Є–≤–∞–љ–Є–µ–Љ –њ–∞–ї—М—Ж–∞ –њ–Њ –њ—А–Њ–±–Є—А–Ї–µ –Є –Њ—В—Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞—В—М.
12. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Б–ї–Є—В—М, –∞ –Њ—Б–∞–і–Њ–Ї —А–∞–Ј—А—Л—Е–ї–Є—В—М –Є –Ј–∞–ї–Є—В—М —В—С–њ–ї—Л–Љ (37¬∞) –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Є–Љ —А–∞—Б—В–≤–Њ—А–Њ–Љ —Е–ї–Њ—А–Є—Б—В–Њ–≥–Њ –Ї–∞–ї–Є—П (0,5%-–љ—Л–є). –Ф–ї—П –њ–Њ–ї—Г—З–µ–љ–Є—П —Е–Њ—А–Њ—И–µ–≥–Њ —А–∞–Ј–±—А–Њ—Б–∞ —Е—А–Њ–Љ–Њ—Б–Њ–Љ –≤ –Љ–µ—В–∞—Д–∞–Ј–љ—Л—Е –њ–ї–∞—Б—В–Є–љ–Ї–∞—Е –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ, —З—В–Њ–±—Л –њ—А–µ–±—Л–≤–∞–љ–Є–µ –Ї–ї–µ—В–Њ–Ї –≤ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ (–≤–Ї–ї—О—З–∞—П —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ) –њ—А–Њ–і–Њ–ї–ґ–∞–ї–Њ—Б—М –љ–µ –±–Њ–ї–µ–µ 16 –Љ–Є–љ.
13. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Б–ї–Є—В—М, –Њ—Б—В–∞–≤–Є—В—М –љ–µ–Љ–љ–Њ–≥–Њ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –љ–∞ –і–љ–µ –њ—А–Њ–±–Є—А–Ї–Є. –Ю—Б–∞–і–Њ–Ї –≤–Ј–±–Њ–ї—В–∞—В—М. –§–Є–Ї—Б–Є—А–Њ–≤–∞—В—М —Б–Љ–µ—Б—М—О –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ —Б –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є (3:1). –•–Њ—А–Њ—И–Њ —Д–Є–Ї—Б–∞—В–Њ—А –њ–Њ–і–∞–≤–∞—В—М –њ–Њ–і –љ–∞–њ–Њ—А–Њ–Љ, –њ—А–Є —Н—В–Њ–Љ —А–∞–Ј–±–Є–≤–∞—О—В—Б—П –≤—Б–µ –Ї—Г—Б–Њ—З–Ї–Є. –Ф–∞—В—М –љ–µ–Љ–љ–Њ–≥–Њ –њ–Њ—Б—В–Њ—П—В—М (3-4 –Љ–Є–љ.) –Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞—В—М.
14. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Г–±—А–∞—В—М –њ–Є–њ–µ—В–Ї–Њ–є, –Њ—Б–∞–і–Њ–Ї —А–µ—Б—Г—Б–њ–µ–љ–Ј–Є—А–Њ–≤–∞—В—М, –≤–ї–Є—В—М –љ–Њ–≤—Г—О –њ–Њ—А—Ж–Є—О —Д–Є–Ї—Б–∞—В–Њ—А–∞ –Є –≤–љ–Њ–≤—М —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞—В—М. –Ґ–∞–Ї –њ–Њ–≤—В–Њ—А–Є—В—М 2-3 —А–∞–Ј–∞. –Т –Є—В–Њ–≥–µ –Њ—Б–∞–і–Њ–Ї –і–Њ–ї–ґ–µ–љ –±—Л—В—М –Њ—З–µ–љ—М –±–µ–ї—Л–Љ. –Х—Б–ї–Є –Њ–љ –µ—Й–µ –Њ–Ї—А–∞—И–µ–љ, —В–Њ –≤–љ–Њ–≤—М —Б–Љ–µ–љ–Є—В—М —Д–Є–Ї—Б–∞—В–Њ—А.
–Ф–∞–ї—М–љ–µ–є—И–µ–µ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –њ—А–Њ–≤–Њ–і–Є—В—Б—П —В–∞–Ї, –Ї–∞–Ї –Њ–њ–Є—Б–∞–љ–Њ –≤ —А–∞–Ј–і–µ–ї–µ ¬Ђ–Ь–Х–Ґ–Ю–Ф –Т–Ђ–°–£–®–Х–Э–Э–Ђ–• –Я–†–Х–Я–Р–†–Р–Ґ–Ю–Т¬ї.
–Ь–Ш–Ъ–†–Ю–Ь–Х–Ґ–Ю–Ф
–Ь–Є–Ї—А–Њ–Љ–µ—В–Њ–і –њ—А–Є–Љ–µ–љ—П–µ—В—Б—П –≤ —В–µ—Е —Б–ї—Г—З–∞—П—Е, –Ї–Њ–≥–і–∞ –≤ —А–∞—Б–њ–Њ—А—П–ґ–µ–љ–Є–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—П –Є–Љ–µ–µ—В—Б—П –љ–µ–±–Њ–ї—М—И–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Ї—А–Њ–≤–Є.
–Т —В—Г–±–µ—А–Ї—Г–ї–Є–љ–Њ–≤—Л–є —И–њ—А–Є—Ж, –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ —Б–Љ–Њ—З–µ–љ–љ—Л–є –≥–µ–њ–∞—А–Є–љ–Њ–Љ, –љ–∞–±–Є—А–∞–µ—В—Б—П –Ї—А–Њ–≤—М –Є–Ј –њ–∞–ї—М—Ж–∞, —Г—Е–∞, —Е–≤–Њ—Б—В–Њ–≤–Њ–є –≤–µ–љ—Л –ґ–Є–≤–Њ—В–љ–Њ–≥–Њ, –ї–Є–±–Њ –Ї–∞–Ї-—В–Њ –Є–љ–∞—З–µ. –≠—В–∞ –Ї—А–Њ–≤—М –≤–≤–Њ–і–Є—В—Б—П –≤ –Ј–∞—А–∞–љ–µ–µ –њ–Њ–і–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–µ —Б—В–µ—А–Є–ї—М–љ—Л–µ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Л–µ –њ—А–Њ–±–Є—А–Ї–Є, —Б–Њ–і–µ—А–ґ–∞—Й–Є–µ 6 –Љ–ї –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і—Л (—Б—А–µ–і–∞ 199 –Є–ї–Є –Ш–≥–ї–∞ —Б –≥–ї—Г—В–∞–Љ–Є–љ–Њ–Љ), 1 –Љ–ї —Б—Л–≤–Њ—А–Њ—В–Ї–Є –Ї—А—Г–њ–љ–Њ–≥–Њ —А–Њ–≥–∞—В–Њ–≥–Њ —Б–Ї–Њ—В–∞ –±–µ–Ј –Ї–Њ–љ—Б–µ—А–≤–∞–љ—В–∞ –Є —Д–Є—В–Њ–≥–µ–Љ–∞–≥–≥–ї—О—В–Є–љ–Є–љ (–Є–Ј —А–∞—Б—З—С—В–∞ 0,2 –Љ–ї –љ–∞ 10 –Љ–ї —Б—А–µ–і—Л). –Ю–±—П–Ј–∞—В–µ–ї—М–љ–Њ –і–Њ–±–∞–≤–ї—П—В—М –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Є, —В–∞–Ї –Ї–∞–Ї –Љ–∞–ї–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Ї—А–Њ–≤–Є –Њ–±—Л—З–љ–Њ –љ–µ —Г–і–∞—С—В—Б—П –≤–Ј—П—В—М —Б—В–µ—А–Є–ї—М–љ–Њ. –°–Њ–і–µ—А–ґ–Є–Љ–Њ–µ –њ—А–Њ–±–Є—А–Ї–Є —В—Й–∞—В–µ–ї—М–љ–Њ –њ–µ—А–µ–Љ–µ—И–Є–≤–∞–µ—В—Б—П –Є –њ—А–Њ–±–Є—А–Ї–Є –њ–Њ–Љ–µ—Й–∞—О—В –њ–Њ–і —Г–≥–ї–Њ–Љ 45¬∞ –≤ —В–µ—А–Љ–Њ—Б—В–∞—В –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞–°. –Ш–љ–Ї—Г–±–Є—А—Г—О—В –Љ–Є–Ї—А–Њ–Ї—Г–ї—М—В—Г—А—Г 3-5 —Б—Г—В–Њ–Ї. –Т–≤–µ–і–µ–љ–Є–µ –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞ –Є –≤—Б—П –њ–Њ—Б–ї–µ–і—Г—О—Й–∞—П –Њ–±—А–∞–±–Њ—В–Ї–∞ –≤–µ–і—Г—В—Б—П —В–∞–Ї –ґ–µ, –Ї–∞–Ї –Є –њ—А–Є –Љ–∞–Ї—А–Њ–Ї—Г–ї—М—В—Г—А–µ (—Б–Љ. –≤—Л—И–µ).
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–Х –Я–†–Х–Я–Р–†–Р–Ґ–Ђ –Ш–Ч –Ъ–£–Ы–ђ–Ґ–£–†–Ђ –≠–Ь–С–†–Ш–Ю–Э–Р–Ы–ђ–Э–Ђ–• –§–Ш–С–†–Ю–С–Ы–Р–°–Ґ–Ю–Т
–≠–Љ–±—А–Є–Њ–љ –Є–Ј–≤–ї–µ–Ї–∞–µ—В—Б—П –њ–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є —Б—В–µ—А–Є–ї—М–љ–Њ –Є –њ–µ—А–µ–љ–Њ—Б–Є—В—Б—П –≤ —А–∞—Б—В–≤–Њ—А –•–µ–љ–Ї—Б–∞, —Б–Њ–і–µ—А–ґ–∞—Й–Є–є –њ–µ–љ–Є—Ж–Є–ї–ї–Є–љ –Є —Б—В—А–µ–њ—В–Њ–Љ–Є—Ж–Є–љ –≤ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є 500-1000 –µ–і/–Љ–ї. –Ь–∞—В–µ—А–Є–∞–ї –Љ–Њ–ґ–µ—В —Е—А–∞–љ–Є—В—М—Б—П –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї–µ –њ—А–Є 4¬∞–° –і–Њ –≤–≤–µ–і–µ–љ–Є—П –µ–≥–Њ –≤ –Ї—Г–ї—М—В—Г—А—Г –≤ —В–µ—З–µ–љ–Є–µ —В—А—С—Е —Б—Г—В–Њ–Ї.
–Я—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –ї—О–±–∞—П —Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ–∞—П —В–Ї–∞–љ—М –њ—А–Є–≥–Њ–і–љ–∞ –і–ї—П –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є—П –Є —Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П. –І–∞—Й–µ –≤—Б–µ–≥–Њ —Ж–Є—В–Њ–≥–µ–љ–µ—В–Є–Ї–∞–Љ–Є –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –Ї—Г–ї—М—В—Г—А–∞ —Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ—Л—Е —Д–Є–±—А–Њ–±–ї–∞—Б—В–Њ–≤.
–Т—Б–µ –њ—А–Њ—Ж–µ–і—Г—А—Л –њ—А–Њ–≤–Њ–і—П—В—Б—П —Б —Б–Њ–±–ї—О–і–µ–љ–Є–µ–Љ –њ—А–∞–≤–Є–ї —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є –≤ –±–Њ–Ї—Б–µ, –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ –Њ–±–ї—Г—З—С–љ–љ–Њ–Љ –±–∞–Ї—В–µ—А–Є—Ж–Є–і–љ—Л–Љ–Є –ї–∞–Љ–њ–∞–Љ–Є –≤ —В–µ—З–µ–љ–Є–µ 1,5 —З–∞—Б.
1. –Т —З–∞—И–Ї–µ –Я–µ—В—А–Є —Б–Њ —Б—В–µ—А–Є–ї—М–љ—Л–Љ —А–∞—Б—В–≤–Њ—А–Њ–Љ –•–µ–љ–Ї—Б–∞ –Њ—В–і–µ–ї–Є—В—М –Њ—В —Н–Љ–±—А–Є–Њ–љ–Њ–≤ –Ї—Г—Б–Њ—З–Ї–Є –Ї–Њ–ґ–љ–Њ–Љ—Л—И–µ—З–љ–Њ–є —В–Ї–∞–љ–Є –Є —В—Й–∞—В–µ–ї—М–љ–Њ –Њ—З–Є—Б—В–Є—В—М –Є—Е –Њ—В —Б–≥—Г—Б—В–Ї–Њ–≤ –Ї—А–Њ–≤–Є.
2. –Ъ—Г—Б–Њ—З–Ї–Є –њ–µ—А–µ–ї–Њ–ґ–Є—В—М –≤–Њ –≤—В–Њ—А—Г—О —З–∞—И–Ї—Г –Я–µ—В—А–Є –Є –њ—А–Њ—Ж–µ–і—Г—А—Г –Њ—З–Є—Б—В–Ї–Є –њ–Њ–≤—В–Њ—А–Є—В—М.
3. –Т—Б–µ –Њ—З–Є—Й–µ–љ–љ—Л–µ –Ї—Г—Б–Њ—З–Ї–Є —Б–Њ–±—А–∞—В—М –≤ –Њ–і–љ–Њ–Љ —Б—В–∞–Ї–∞–љ–µ —Б–Њ —Б–≤–µ–ґ–Є–Љ —А–∞—Б—В–≤–Њ—А–Њ–Љ –•–µ–љ–Ї—Б–∞ –Є —В—Й–∞—В–µ–ї—М–љ–Њ –Є—Е –Є–Ј–Љ–µ–ї—М—З–Є—В—М –љ–Њ–ґ–љ–Є—Ж–∞–Љ–Є –і–Њ –Ї–Њ–љ—Б–Є—Б—В–µ–љ—Ж–Є–Є —Д–∞—А—И–∞.
4. –Ь–∞—В–µ—А–Є–∞–ї –њ–µ—А–µ–љ–µ—Б—В–Є –≤ —Б—В–µ—А–Є–ї—М–љ—Г—О –Ї–Њ–ї–±—Г, —В—Г–і–∞ –љ–∞–ї–Є—В—М —Е–Њ–ї–Њ–і–љ—Л–є 0,25%-–љ—Л–є —А–∞—Б—В–≤–Њ—А —В—А–Є–њ—Б–Є–љ–∞, –Ї–Њ–ї–±—Г –њ–ї–Њ—В–љ–Њ –Ј–∞–Ї—А—Л—В—М —Б—В–µ—А–Є–ї—М–љ–Њ–є —А–µ–Ј–Є–љ–Њ–≤–Њ–є –њ—А–Њ–±–Ї–Њ–є –Є–ї–Є —Д–Њ–ї—М–≥–Њ–є –Є –њ–Њ—Б—В–∞–≤–Є—В—М –≤ —В–µ—А–Љ–Њ—Б—В–∞—В (–Є–ї–Є –≤–Њ–і—П–љ—Г—О –±–∞–љ—О) –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞ –љ–∞ 1 —З–∞—Б.
5. –°–Њ–і–µ—А–ґ–Є–Љ–Њ–µ –Ї–Њ–ї–±—Л –њ–µ—А–µ–ї–Є—В—М –≤ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Л–µ –њ—А–Њ–±–Є—А–Ї–Є –Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞—В—М 5 –Љ–Є–љ. –њ—А–Є 1000 –Њ–±/–Љ–Є–љ.
6. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ–∞—П –ґ–Є–і–Ї–Њ—Б—В—М —Б–ї–Є–≤–∞–µ—В—Б—П, –∞ –Њ—Б–∞–і–Њ–Ї —Б –љ–µ–±–Њ–ї—М—И–Є–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ–Љ –Њ—Б—В–∞–≤—И–µ–є—Б—П –ґ–Є–і–Ї–Њ—Б—В–Є –≤—Л–ї–Є–≤–∞–µ—В—Б—П –≤ –љ–µ–±–Њ–ї—М—И—Г—О –њ–ї–Њ—Б–Ї–Њ–і–Њ–љ–љ—Г—О –Ї–Њ–ї–±—Г, –Ї—Г–і–∞ –і–Њ–±–∞–≤–ї—П–µ—В—Б—П –њ–Є—В–∞—В–µ–ї—М–љ–∞—П —Б–Љ–µ—Б—М, —Б–Њ–і–µ—А–ґ–∞—Й–∞—П 50% —Б—А–µ–і—Л –Ш–≥–ї–∞ —Б –≥–ї—Г—В–∞–Љ–Є–љ–Њ–Љ, 30% –≥–Є–і—А–Њ–ї–Є–Ј–∞—В–∞ –ї–∞–Ї—В–∞–ї—М–±—Г–Љ–Є–љ–∞ –Є 20% —Б—Л–≤–Њ—А–Њ—В–Ї–Є –Ї—А—Г–њ–љ–Њ–≥–Њ —А–Њ–≥–∞—В–Њ–≥–Њ —Б–Ї–Њ—В–∞.
7. –≠—В–∞ –≤–Ј–≤–µ—Б—М —Б—В–∞–≤–Є—В—Б—П –љ–∞ –Љ–∞–≥–љ–Є—В–љ—Г—О –Љ–µ—И–∞–ї–Ї—Г –љ–∞ 10 –Љ–Є–љ.
8. –§–Є–ї—М—В—А—Г–µ—В—Б—П –≤ –Љ–µ—А–љ—Л–є —Д–ї–∞–Ї–Њ–љ —З–µ—А–µ–Ј —Б—В–µ–Ї–ї—П–љ–љ—Л–є —Д–Є–ї—М—В—А G1 (–Љ–Њ–ґ–љ–Њ —З–µ—А–µ–Ј –≤–Њ—А–Њ–љ–Ї—Г —Б –љ–µ—Б–Ї–Њ–ї—М–Ї–Є–Љ–Є —Б–ї–Њ—П–Љ–Є –Љ–∞—А–ї–Є –≤–Љ–µ—Б—В–Њ —Д–Є–ї—М—В—А–∞). –Т –Њ—Б—В–∞–≤—И–Є–є—Б—П –љ–∞ –і–љ–µ –Њ—Б–∞–і–Њ–Ї –≤–љ–Њ–≤—М –љ–∞–ї–Є–≤–∞–µ—В—Б—П –њ–Є—В–∞-—В–µ–ї—М–љ–∞—П —Б—А–µ–і–∞ –Є –Ї–Њ–ї–±–∞ –≤–љ–Њ–≤—М —Б—В–∞–≤–Є—В—Б—П –љ–∞ –Љ–∞–≥–љ–Є—В–љ—Г—О –Љ–µ—И–∞–ї–Ї—Г –љ–∞ 10 –Љ–Є–љ., –њ–Њ—Б–ї–µ —З–µ–≥–Њ –ґ–Є–і–Ї–Њ—Б—В—М —Д–Є–ї—М—В—А—Г–µ—В—Б—П –≤ —В–Њ—В –ґ–µ –Љ–µ—А–љ—Л–є —Д–ї–∞–Ї–Њ–љ.
9. –°—В–µ—А–Є–ї—М–љ–Њ–є –њ–Є–њ–µ—В–Ї–Њ–є –≤–Ј—П—В—М –Ї–∞–њ–ї—О —Б—Г—Б–њ–µ–љ–Ј–Є–Є –Є –≤ –Ї–∞–Љ–µ—А–µ –У–Њ—А—П–µ–≤–∞ –њ–Њ–і—Б—З–Є—В–∞—В—М –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Ї–ї–µ—В–Њ–Ї, –њ—А–Є—Е–Њ–і—П—Й–Є—Е—Б—П –≤ —Б—А–µ–і–љ–µ–Љ –љ–∞ –Њ–і–Є–љ –Ї–≤–∞–і—А–∞—В –Є–Ј –і–µ—Б—П—В–Є –њ—А–Њ—Б—З–Є—В–∞–љ–љ—Л—Е. –Ф–∞–ї–µ–µ –≤—Л—Б—З–Є—В—Л–≤–∞—О—В, –≤–Њ —Б–Ї–Њ–ї—М–Ї–Њ —А–∞–Ј —Б–ї–µ–і—Г–µ—В —А–∞–Ј–±–∞–≤–Є—В—М —Б—Г—Б–њ–µ–љ–Ј–Є—О —Б—А–µ–і–Њ–є, —З—В–Њ–±—Л –≤ –Є—В–Њ–≥–µ –љ–∞ –Њ–і–Є–љ –Ї–≤–∞–і—А–∞—В –Ї–∞–Љ–µ—А—Л –У–Њ—А—П–µ–≤–∞ –њ—А–Є—Е–Њ–і–Є–ї–∞—Б—М —В–Њ–ї—М–Ї–Њ –Њ–і–љ–∞ –Ї–ї–µ—В–Ї–∞.
10. –Т–Ј–≤–µ—Б—М –≤—Л–ї–Є–≤–∞–µ—В—Б—П –≤ –Љ–∞—В—А–∞—Б –Є–ї–Є –њ–ї–Њ—Б–Ї–Њ–і–Њ–љ–љ—Л–µ –Ї–Њ–ї–±—Л, –Ј–∞–Ї—А—Л–≤–∞–µ—В—Б—П —Б—В–µ—А–Є–ї—М–љ–Њ–є —А–µ–Ј–Є–љ–Њ–≤–Њ–є –њ—А–Њ–±–Ї–Њ–є –Є —Б–≤–µ—А—Е—Г —Д–Њ–ї—М–≥–Њ–є –Є –њ–Њ–Љ–µ—Й–∞–µ—В—Б—П –≤ —В–µ—А–Љ–Њ—Б—В–∞—В –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞ –љ–∞ 48-72 —З–∞—Б–∞. –Ь–Є—В–Њ—В–Є—З–µ—Б–Ї–Є–µ –і–µ–ї–µ–љ–Є—П –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Ї–ї–µ—В–Њ–Ї –љ–∞—З–Є–љ–∞—О—В—Б—П —З–µ—А–µ–Ј 48 —З–∞—Б. –Њ—В –љ–∞—З–∞–ї–∞ –Є–љ–Ї—Г–±–∞—Ж–Є–Є –Є –њ–Њ—З—В–Є –њ—А–µ–Ї—А–∞—Й–∞—О—В—Б—П —З–µ—А–µ–Ј 72 —З–∞—Б–∞.
11. –Ч–∞ 1-2 —З–∞—Б–∞ –і–Њ –Њ–Ї–Њ–љ—З–∞–љ–Є—П –Є–љ–Ї—Г–±–∞—Ж–Є–Є –і–Њ–±–∞–≤–ї—П–µ—В—Б—П –њ–Њ–і–Њ–≥—А–µ—В—Л–є –≤–Њ–і–љ—Л–є —А–∞–±–Њ—З–Є–є —А–∞—Б—В–≤–Њ—А –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞ –Є–Ј —А–∞—Б—З—С—В–∞ 0,5 –≥–∞–Љ–Љ—Л –љ–∞ 1 –Љ–ї —Б—А–µ–і—Л (—А–∞–±–Њ—З–Є–є —А–∞—Б—В–≤–Њ—А –Ї–Њ–ї—Е–Є—Ж–Є–љ–∞ —Б–Њ–і–µ—А–ґ–Є—В 20 –≥–∞–Љ–Љ –≤ 1 –Љ–ї). –Т –і–∞–ї—М–љ–µ–є—И–Є—Е –Њ–њ–µ—А–∞—Ж–Є—П—Е —Б–Њ–±–ї—О–і–µ–љ–Є—П —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є –љ–µ —В—А–µ–±—Г–µ—В—Б—П.
12. –Я–Њ –Њ–Ї–Њ–љ—З–∞–љ–Є–Є –Є–љ–Ї—Г–±–∞—Ж–Є–Є —Б–ї–Є—В—М —Б—А–µ–і—Г –Є –љ–∞–ї–Є—В—М –љ–µ–Љ–љ–Њ–≥–Њ —Е–Њ–ї–Њ–і–љ–Њ–≥–Њ —В—А–Є–њ—Б–Є–љ–∞, –Ї–Њ—В–Њ—А—Л–є —В—Г—В –ґ–µ –≤—Л–ї–Є—В—М. –Я–Њ–і–Њ–ґ–і–∞—В—М –љ–µ–Љ–љ–Њ–≥–Њ, —З—В–Њ–±—Л –Ї–ї–µ—В–Ї–Є –љ–∞—З–∞–ї–Є —Б–њ–Њ–ї–Ј–∞—В—М —Б–Њ —Б—В–µ–Ї–ї–∞, –Є –≤–ї–Є—В—М —А–∞—Б—В–≤–Њ—А –•–µ–љ–Ї—Б–∞. –Ґ—Й–∞—В–µ–ї—М–љ–Њ –≤—Б—В—А—П—Е–љ—Г—В—М –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —А–∞–Ј, —З—В–Њ–±—Л —Б–Љ—Л—В—М –≤—Б–µ –Ї–ї–µ—В–Ї–Є.
13. –Т–Ј–≤–µ—Б—М —А–∞–Ј–ї–Є—В—М –њ–Њ —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Л–Љ –њ—А–Њ–±–Є—А–Ї–∞–Љ –Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞—В—М 5-8 –Љ–Є–љ. –њ—А–Є 1000 –Њ–±/–Љ–Є–љ.
14. –°–ї–Є—В—М –Є–ї–Є –Њ—В—Б–Њ—Б–∞—В—М –љ–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М, –Њ—Б–∞–і–Њ–Ї –≤–Ј–±–Њ–ї—В–∞—В—М –њ–Њ—Б—В—Г–Ї–Є–≤–∞–љ–Є–µ–Љ –њ–∞–ї—М—Ж–∞ –Є –љ–∞–ї–Є—В—М —В—С–њ–ї—Л–є (37¬∞–°) —А–∞—Б—В–≤–Њ—А 1%-–љ–Њ–≥–Њ —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П (—В—А–µ—Е–Ј–∞–Љ–µ—Й–µ–љ–љ—Л–є –ї–Є–Љ–Њ–љ–љ–Њ–Ї–Є—Б–ї—Л–є –љ–∞—В—А–Є–є).
15. –°–Њ–і–µ—А–ґ–Є–Љ–Њ–µ –њ—А–Њ–±–Є—А–Њ–Ї –≤–Ј–±–Њ–ї—В–∞—В—М –Є –њ–Њ—Б—В–∞–≤–Є—В—М –≤ —В–µ—А–Љ–Њ—Б—В–∞—В –Є–ї–Є –≤–Њ–і—П–љ—Г—О –±–∞–љ—О –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 37¬∞ –љ–∞ 20-30 –Љ–Є–љ. –Я–Њ –Њ–Ї–Њ–љ—З–∞–љ–Є–Є –Є–љ–Ї—Г–±–∞—Ж–Є–Є –њ—А–Њ–±–Є—А–Ї–Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А—Г—О—В.
16. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Г–і–∞–ї—П—О—В, –∞ –Њ—Б–∞–і–Њ–Ї —Б –љ–µ–±–Њ–ї—М—И–Є–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ–Љ –Њ—Б—В–∞–≤—И–µ–є—Б—П –ґ–Є–і–Ї–Њ—Б—В–Є —А–∞–Ј–±–Є–≤–∞—О—В –ї—С–≥–Ї–Є–Љ –њ–Њ—Б—В—Г–Ї–Є–≤–∞–љ–Є–µ–Љ –њ–∞–ї—М—Ж–µ–≤ –њ–Њ –њ—А–Њ–±–Є—А–Ї–µ –Є –љ–∞–ї–Є–≤–∞—О—В –Њ—Е–ї–∞–ґ–і—С–љ–љ—Л–є —Д–Є–Ї—Б–∞—В–Њ—А, —Б–Њ—Б—В–Њ—П—Й–Є–є –Є–Ј –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л (3:1).
17. –Я–Њ—Б–ї–µ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є—П –љ–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Г–і–∞–ї—П—О—В, –Њ—Б–∞–і–Њ–Ї —А–∞–Ј–±–Є–≤–∞—О—В –Є –Ј–∞–ї–Є–≤–∞—О—В –љ–Њ–≤—Л–Љ —Д–Є–Ї—Б–∞—В–Њ—А–Њ–Љ, —Б–Њ—Б—В–Њ—П—Й–Є–Љ –Є–Ј –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л, –љ–Њ –≤ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–Є 2:1. –Ю–±—Й–∞—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Д–Є–Ї—Б–∞—Ж–Є–Є 1 —З–∞—Б –Є–ї–Є –±–Њ–ї–µ–µ.
–Ф–∞–ї—М–љ–µ–є—И–µ–µ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –≤–µ–і—С—В—Б—П –њ–Њ –Њ–±—Л—З–љ–Њ–є –Љ–µ—В–Њ–і–Є–Ї–µ ¬Ђ–≤—Л—Б—Г—И–µ–љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤¬ї.
–Я–†–Ш–У–Ю–Ґ–Ю–Т–Ы–Х–Э–Ш–Х –Я–†–Х–Я–Р–†–Р–Ґ–Ю–Т –Ь–Х–Щ–Ю–Ґ–Ш–І–Х–°–Ъ–Ш–• –•–†–Ю–Ь–Ю–°–Ю–Ь
–Т –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П —А–∞–Ј—А–∞–±–Њ—В–∞–љ—Л –Є —Г—Б–њ–µ—И–љ–Њ –њ—А–Є–Љ–µ–љ—П—О—В—Б—П –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Љ–µ—В–Њ–і–Њ–≤ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Ј —Б–µ–Љ–µ–љ–љ–Є–Ї–Њ–≤ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Ь–µ–є–Њ—В–Є—З–µ—Б–Ї–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л –Є—Б—Б–ї–µ–і—Г—О—В –Ї–∞–Ї –љ–∞ –і–∞–≤–ї–µ–љ—Л—Е —В–Њ—В–∞–ї—М–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–∞—Е –њ–Њ–ї–Њ–≤—Л—Е –ґ–µ–ї—С–Ј, —В–∞–Ї –Є –љ–∞ –њ—А–µ–њ–∞—А–∞—В–∞—Е, –њ–Њ–ї—Г—З–µ–љ–љ—Л—Е –њ—Г—В—С–Љ –≤—Л—Б—Г—И–Є–≤–∞–љ–Є—П –≤–Ј–≤–µ—Б–Є –Ї–ї–µ—В–Њ–Ї, –љ–∞–љ–µ—Б—С–љ–љ—Л—Е –љ–∞ –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ.
–Т –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П —П–≤–љ–Њ–µ –њ—А–µ–і–њ–Њ—З—В–µ–љ–Є–µ –Њ—В–і–∞—О—В –≤—В–Њ—А–Њ–Љ—Г –Љ–µ—В–Њ–і—Г, —В–∞–Ї –Ї–∞–Ї –Љ–µ—В–Њ–і –і–∞–≤–ї–µ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Љ–µ–µ—В —А—П–і —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л—Е –љ–µ–і–Њ—Б—В–∞—В–Ї–Њ–≤. –Э–Є–ґ–µ –Љ—Л –њ—А–Є–≤–Њ–і–Є–Љ –і–≤–µ –љ–∞–Є–±–Њ–ї–µ–µ –њ–Њ–њ—Г–ї—П—А–љ—Л–µ –Љ–µ—В–Њ–і–Є–Ї–Є, –њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ–µ —А–∞–Ј–ї–Є—З–Є–µ –Ї–Њ—В–Њ—А—Л—Е –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤ —Б–њ–Њ—Б–Њ–±–µ –≤—Л–і–µ–ї–µ–љ–Є—П –Ї–ї–µ—В–Њ–Ї –Є–Ј –≥–µ–љ–µ—А–∞—В–Є–≤–љ–Њ–є —В–Ї–∞–љ–Є.
–С–Њ–ї—М—И–∞—П —З–∞—Б—В—М –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–Њ –Љ–µ–є–Њ–Ј—Г —Г –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –≤—Л–њ–Њ–ї–љ–µ–љ–љ—Л—Е –≤ –њ–Њ—Б–ї–µ–і–љ–Є–µ –≥–Њ–і—Л, –±–∞–Ј–Є—А—Г–µ—В—Б—П –љ–∞ –њ—А–µ–њ–∞—А–∞—В–∞—Е, –њ–Њ–ї—Г—З–µ–љ–љ—Л—Е –њ–Њ –Љ–µ—В–Њ–і–Є–Ї–µ –Ш–≤–µ–љ—Б–∞ –Є —Б–Њ–∞–≤—В–Њ—А–Њ–≤ (Evans et al., 1964).
1. –°–µ–Љ–µ–љ–љ–Є–Ї–Є –њ–Њ–Љ–µ—Й–∞—О—В –≤ –љ–µ–±–Њ–ї—М—И–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ 2,2%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —В—А—С—Е–Ј–∞–Љ–µ—Й—С–љ–љ–Њ–≥–Њ —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П –њ—А–Є –Ї–Њ–Љ–љ–∞—В–љ–Њ–є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ. –Э–∞–і—А–µ–Ј–∞—О—В —В—Г–љ–Є–Ї—Г –Є —Б–µ–Љ–µ–љ–љ—Л–µ –Ї–∞–љ–∞–ї—М—Ж—Л –≤—Л–њ—Г—Б–Ї–∞—О—В –≤ —А–∞—Б—В–≤–Њ—А –і–ї—П —Г–і–∞–ї–µ–љ–Є—П —Б–Њ–њ—Г—В—Б—В–≤—Г—О—Й–µ–≥–Њ –ґ–Є—А–∞ –Є –Є–љ—В–µ—А—Б—В–Є—Ж–Є–∞–ї—М–љ–Њ–є —В–Ї–∞–љ–Є. –Я–Њ—Б–ї–µ —Н—В–Є—Е –Њ–њ–µ—А–∞—Ж–Є–є —Б–µ–Љ–µ–љ–љ—Л–µ –Ї–∞–љ–∞–ї—М—Ж—Л –њ–µ—А–µ–љ–Њ—Б—П—В –≤ —Б–≤–µ–ґ–Є–є —А–∞—Б—В–≤–Њ—А —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П —В–∞–Ї–Њ–є –ґ–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є.
2. –Ф–µ—А–ґ–∞ –Љ–∞—Б—Б—Г —Б–µ–Љ–µ–љ–љ—Л—Е –Ї–∞–љ–∞–ї—М—Ж–µ–≤ –≥–ї–∞–Ј–љ—Л–Љ –њ–Є–љ—Ж–µ—В–Њ–Љ, –і—А—Г–≥–Є–Љ –њ–Є–љ—Ж–µ—В–Њ–Љ –Њ—Б—В–Њ—А–Њ–ґ–љ–Њ, —З—В–Њ–±—Л –љ–µ –њ–Њ—А–≤–∞—В—М –Ї–∞–љ–∞–ї—М—Ж—Л, –≤—Л–і–∞–≤–ї–Є–≤–∞—О—В —Б–Њ–і–µ—А–ґ–Є–Љ–Њ–µ –Ї–∞–љ–∞–ї—М—Ж–µ–≤ –≤ —А–∞—Б—В–≤–Њ—А. –Я–Њ–ї—Г—З–µ–љ–љ—Г—О –Ї–ї–µ—В–Њ—З–љ—Г—О —Б—Г—Б–њ–µ–љ–Ј–Є—О –њ–µ—А–µ–љ–Њ—Б—П—В –≤ –њ—А–Њ–±–Є—А–Ї—Г, –і–Њ–±–∞–≤–ї—П—О—В –љ–µ–Љ–љ–Њ–≥–Њ (–Њ–Ї–Њ–ї–Њ 3 –Љ–ї) —Б–≤–µ–ґ–µ–≥–Њ —Ж–Є—В—А–∞—В–∞ –Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А—Г—О—В 30 —Б–µ–Ї. –њ—А–Є 500 –Њ–±/–Љ–Є–љ. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М –њ–µ—А–µ–ї–Є–≤–∞—О—В –≤ –љ–Њ–≤—Г—О —Ж–µ–љ—В—А–Є—Д—Г–ґ–љ—Г—О –њ—А–Њ–±–Є—А–Ї—Г –Є —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А—Г—О—В 5 –Љ–Є–љ. –њ—А–Є 500 –Њ–±/–Љ–Є–љ. –Я—А–Є —В–∞–Ї–Њ–Љ —А–µ–ґ–Є–Љ–µ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є—П —Б–њ–µ—А–Љ–Є–Є –Њ—Б—В–∞—О—В—Б—П –≤ —Б—Г—Б–њ–µ–љ–Ј–Є–Є, –∞ —Б–њ–µ—А–Љ–∞—В–Њ—Ж–Є—В—Л –Њ—Б–∞–ґ–і–∞—О—В—Б—П –љ–∞ –і–љ–Њ –њ—А–Њ–±–Є—А–Ї–Є.
3. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М —Б–ї–Є–≤–∞—О—В, –∞ –Њ—Б–∞–і–Њ–Ї —А–µ—Б—Г—Б–њ–µ–љ–Ј–Є—А—Г—О—В –≤ 3 –Љ–ї 1%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П –ї—С–≥–Ї–Є–Љ –њ–Њ—Б—В—Г–Ї–Є–≤–∞–љ–Є–µ–Љ –њ–∞–ї—М—Ж–µ–≤ –њ–Њ —Б—В–µ–љ–Ї–µ –њ—А–Њ–±–Є—А–Ї–Є. –Т —В–µ—З–µ–љ–Є–µ 7-12 –Љ–Є–љ. (–≤ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В —В–Њ–≥–Њ, —Б —Б–µ–Љ–µ–љ–љ–Є–Ї–∞–Љ–Є –Ї–∞–Ї–Њ–≥–Њ –≤–Є–і–∞ –ґ–Є–≤–Њ—В–љ–Њ–≥–Њ –Є–Љ–µ—О—В –і–µ–ї–Њ) –Ї–ї–µ—В–Ї–Є –≤—Л–і–µ—А–ґ–Є–≤–∞—О—В—Б—П –≤ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ –њ—А–Є –Ї–Њ–Љ–љ–∞—В–љ–Њ–є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ (–і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–є –Њ–±—А–∞–±–Њ—В–Ї–Є –њ–Њ–і–±–Є—А–∞–µ—В—Б—П —Н–Љ–њ–Є—А–Є—З–µ—Б–Ї–Є). –Ґ–∞–Ї, –њ–Њ –і–∞–љ–љ—Л–Љ –Р.–Ш. –Ъ–Њ–Ј–ї–Њ–≤—Б–Ї–Њ–≥–Њ, –њ—А–µ–±—Л–≤–∞–љ–Є–µ –Ї–ї–µ—В–Њ–Ї –≤ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ –≤ —В–µ—З–µ–љ–Є–µ 12 –Љ–Є–љ. –њ–Њ–ї–љ–Њ—Б—В—М—О —А–∞–Ј—А—Г—И–∞–µ—В —Б–њ–µ—А–Љ–∞—В–Њ—Ж–Є—В—Л —Г –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –Є –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л–Љ –і–ї—П —Н—В–Њ–≥–Њ –≤–Є–і–∞ —П–≤–ї—П–µ—В—Б—П –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М—О 7вАУ10 –Љ–Є–љ.
4. –Ъ–ї–µ—В–Ї–Є –Њ—Б–∞–ґ–і–∞—О—В 5-–Љ–Є–љ—Г—В–љ—Л–Љ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ–Љ –њ—А–Є 500 –Њ–±/–Љ–Є–љ. –Э–∞–і–Њ—Б–∞–і–Њ—З–љ—Г—О –ґ–Є–і–Ї–Њ—Б—В—М –Њ—Б—В–Њ—А–Њ–ґ–љ–Њ —Г–і–∞–ї—П—О—В, –∞ –Њ—Б–∞–і–Њ–Ї —Б—А–∞–Ј—Г –ґ–µ —А–µ—Б—Г—Б–њ–µ–љ–Ј–Є—А—Г—О—В –ї—С–≥–Ї–Є–Љ –≤—Б—В—А—П—Е–Є–≤–∞–љ–Є–µ–Љ –њ—А–Њ–±–Є—А–Ї–Є.
5. –Ю—Б—В–Њ—А–Њ–ґ–љ–Њ, –њ–Њ –Ї–∞–њ–ї—П–Љ, –≤—Б–µ –≤—А–µ–Љ—П, –≤–Ј–±–∞–ї—В—Л–≤–∞—П –Є –њ–Њ—Б—В—Г–Ї–Є–≤–∞—П –њ–∞–ї—М—Ж–µ–Љ –њ–Њ –њ—А–Њ–±–Є—А–Ї–µ, –≤–ї–Є–≤–∞—О—В —Д–Є–Ї—Б–∞—В–Њ—А, —Б–Њ—Б—В–Њ—П—Й–Є–є –Є–Ј 1 —З–∞—Б—В–Є –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л, 3 —З–∞—Б—В–µ–є –Љ–µ—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є–ї–Є –∞–±—Б–Њ–ї—О—В–љ–Њ–≥–Њ —Н—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є —Е–ї–Њ—А–Њ—Д–Њ—А–Љ–∞ (1 —З–∞—Б—В—М –љ–∞ 40 —З–∞—Б—В–µ–є —Д–Є–Ї—Б–∞—В–Њ—А–∞). –Ъ–Њ–ї–Є—З–µ—Б—В–≤–Њ —Д–Є–Ї—Б–∞—В–Њ—А–∞ –Њ–Ї–Њ–ї–Њ 2-3 –Љ–ї.
6. –Я–Њ—Б–ї–µ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є—П –Њ—Б–∞–і–Њ–Ї —А–µ—Б—Г—Б–њ–µ–љ–Ј–Є—А—Г—О—В –≤ —Б–≤–µ–ґ–µ–Љ —Д–Є–Ї—Б–∞—В–Њ—А–µ –Є –Њ—Б—В–∞–≤–ї—П—О—В –≤ –љ—С–Љ –љ–∞ 10 –Љ–Є–љ. –Ч–∞—В–µ–Љ —Б–ї–µ–і—Г–µ—В –µ—Й—С –Њ–і–љ–Њ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ –Є –Ј–∞–Љ–µ–љ–∞ —Д–Є–Ї—Б–∞—В–Њ—А–∞.
7. –Э–∞ —Б–Њ–≥—А–µ—В–Њ–µ –њ–Њ–і –ї–∞–Љ–њ–Њ–є –∞–±—Б–Њ–ї—О—В–љ–Њ —З–Є—Б—В–Њ–µ –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ –љ–∞–Ї–∞–њ–∞—В—М 2-3 –Ї–∞–њ–ї–Є —Б—Г—Б–њ–µ–љ–Ј–Є–Є –Є –њ–Њ—Б–ї–µ –њ–Њ–ї–љ–Њ–≥–Њ —А–∞—Б—В–µ–Ї–∞–љ–Є—П –Ї–∞–ґ–і–Њ–є –Ї–∞–њ–ї–Є –њ–Њ–і—Г—В—М –љ–∞ —Б—В–µ–Ї–ї–Њ. –Ф–∞—В—М –њ—А–µ–њ–∞—А–∞—В—Г –њ–Њ–і—Б–Њ—Е–љ—Г—В—М. –≠—В–∞ –Љ–µ—В–Њ–і–Є–Ї–∞ –і–∞—С—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –њ–Њ–ї—Г—З–∞—В—М —Е–Њ—А–Њ—И–Є–µ –њ—А–µ–њ–∞—А–∞—В—Л –Љ–µ–є–Њ—В–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Є–Ј —Б–µ–Љ–µ–љ–љ–Є–Ї–Њ–≤ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –Њ–і–љ–∞–Ї–Њ –љ–µ –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Є—Б—Б–ї–µ–і–Њ–≤–∞—В—М –љ–µ–Ї–Њ—В–Њ—А—Л–µ —Б—В–∞–і–Є–Є –Љ–µ–є–Њ–Ј–∞, –∞ —В–∞–Ї–ґ–µ –Љ–µ—В–∞—Д–∞–Ј—Л. –Т–Њ –≤—А–µ–Љ—П –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П –Є–Ј –љ–µ—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ–љ—Л—Е —Б–µ–Љ–µ–љ–љ—Л—Е –Ї–∞–љ–∞–ї—М—Ж–µ–≤ –≤–Ј–≤–µ—Б–Є –Є–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ї–ї–µ—В–Њ–Ї –Є –њ–Њ–і –і–µ–є—Б—В–≤–Є–µ–Љ –≥–Є–њ–Њ—В–Њ–љ–Є–Є —В–µ—А—П–µ—В—Б—П –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ —Б–њ–µ—А–Љ–∞—В–Њ–≥–Њ–љ–Є–є –љ–∞ —Б—В–∞–і–Є–Є –Љ–µ—В–∞—Д–∞–Ј—Л, –∞ –њ—А–Є –љ–µ–Њ–і–љ–Њ–Ї—А–∞—В–љ–Њ–Љ —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–Є –Є —Б–Љ–µ–љ–µ —Д–Є–Ї—Б–∞—В–Њ—А–∞ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –њ–Њ—В–µ—А—П —Б–њ–µ—А–Љ–∞—В–Њ—Ж–Є—В–Њ–≤ –љ–∞ —Б—В–∞–і–Є–Є –Љ–µ—В–∞—Д–∞–Ј—Л 2-–≥–Њ –і–µ–ї–µ–љ–Є—П —Б–Њ–Ј—А–µ–≤–∞–љ–Є—П (–Ф—Л–±–∞–љ, 1970).
–Я—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–∞—П –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—М –Љ–µ—В–Њ–і–Є–Ї–Є, –Њ–њ–Є—Б—Л–≤–∞–µ–Љ–Њ–є –љ–Є–ґ–µ (Meredith, 1969), —Б–Њ—Б—В–Њ–Є—В –≤ –Є–љ–Њ–є –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В–Є –Њ–±—А–∞–±–Њ—В–Ї–Є —В–Ї–∞–љ–Є. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –Њ–Ї–∞–Ј—Л–≤–∞—О—В—Б—П –Є–Ј–ї–Є—И–љ–Є–Љ–Є —В–∞–Ї–Є–µ –Њ–њ–µ—А–∞—Ж–Є–Є, –Ї–∞–Ї –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є–µ –Є–Ј –љ–µ—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ–љ—Л—Е —Б–µ–Љ–µ–љ–љ—Л—Е –Ї–∞–љ–∞–ї—М—Ж–µ–≤ –≤–Ј–≤–µ—Б–Є –Є–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ї–ї–µ—В–Њ–Ї, —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–љ–Є–µ, —З—В–Њ —Г—Б—В—А–∞–љ—П–µ—В –њ–Њ—В–µ—А–Є –Ї–ї–µ—В–Њ–Ї –≤ —Е–Њ–і–µ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П –њ—А–µ–њ–∞—А–∞—В–Њ–≤.
1. –Ю–Ї–Њ–ї–Њ 100 –Љ–≥ —Б–µ–Љ–µ–љ–љ–Є–Ї–∞ –њ–Њ–Љ–µ—Й–∞—О—В –≤ —Б–≤–µ–ґ–µ–њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–є 1%-–љ—Л–є —А–∞—Б—В–≤–Њ—А —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П, –Њ—В–Љ—Л–≤–∞—О—В –ґ–Є—А –Є –Є–љ—В–µ—А—Б—В–Є—Ж–Є–∞–ї—М–љ—Г—О —В–Ї–∞–љ—М –Є —Г–і–∞–ї—П—О—В —В—Г–љ–Є–Ї—Г.
2. –Ю—В–Љ—Л—В—Л–µ –Ї–∞–љ–∞–ї—М—Ж—Л –њ–µ—А–µ–љ–Њ—Б—П—В –≤ —Б–≤–µ–ґ–Є–є 1%-–љ—Л–є —А–∞—Б—В–≤–Њ—А —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П –Є –Њ—Б—В–Њ—А–Њ–ґ–љ–Њ —А–∞—Б–њ—Г—В—Л–≤–∞—О—В, —З—В–Њ–±—Л –Њ–±–µ—Б–њ–µ—З–Є—В—М —Е–Њ—А–Њ—И–µ–µ –њ—А–Њ–љ–Є–Ї–љ–Њ–≤–µ–љ–Є–µ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞. –Э–µ–Њ–±—Е–Њ–і–Є–Љ–Њ —Б–ї–µ–і–Є—В—М, —З—В–Њ–±—Л –љ–µ –њ—А–Њ–Є–Ј–Њ—И–ї–Њ —А–∞–Ј—А—Л–≤–∞ –Ї–∞–љ–∞–ї—М—Ж–µ–≤.
3. –Я–Њ—Б–ї–µ —Н—В–Њ–є –Њ–±—А–∞–±–Њ—В–Ї–Є –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –њ–Њ–Љ—Г—В–љ–µ–љ–Є–µ —А–∞—Б—В–≤–Њ—А–∞ —Ж–Є—В—А–∞—В–∞ –Є –Љ–∞—В–µ—А–Є–∞–ї —Б–ї–µ–і—Г–µ—В –њ–µ—А–µ–љ–µ—Б—В–Є –і–ї—П –і–∞–ї—М–љ–µ–є—И–µ–є –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–є –Њ–±—А–∞–±–Њ—В–Ї–Є –≤ —В—А–µ—В—М—О —Б–Љ–µ–љ—Г 1%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П. –Ю–±—К–µ–Љ —А–∞—Б—В–≤–Њ—А–∞ —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П –і–Њ–ї–ґ–µ–љ –њ—А–Є–Љ–µ—А–љ–Њ –≤ 20 —А–∞–Ј –њ—А–µ–≤—Л—И–∞—В—М –Њ–±—К–µ–Љ —В–Ї–∞–љ–Є.
4. –Ю–±—Й–µ–µ –≤—А–µ–Љ—П –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–є –Њ–±—А–∞–±–Њ—В–Ї–Є –≤–∞—А—М–Є—А—Г–µ—В –і–ї—П —А–∞–Ј–љ—Л—Е –≤–Є–і–Њ–≤ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Ф–ї—П —Б–µ–Љ–µ–љ–љ—Л—Е –Ї–∞–љ–∞–ї—М—Ж–µ–≤ –Ї—А—Л—Б, —Б–Є—А–Є–є—Б–Ї–Є—Е —Е–Њ–Љ—П—З–Ї–Њ–≤ –Є –Ї—А–Њ–ї–Є–Ї–Њ–≤ –і–Њ—Б—В–∞—В–Њ—З–љ–Њ –≤–Њ—Б—М–Љ–Є–Љ–Є–љ—Г—В–љ–Њ–≥–Њ –њ—А–µ–±—Л–≤–∞–љ–Є—П –≤ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ. –Ф–ї—П —В–Ї–∞–љ–µ–є –Љ–Њ—А—Б–Ї–Њ–є —Б–≤–Є–љ–Ї–Є –Є –±–µ–ї–Њ–є –Љ—Л—И–Є –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В—Б—П –і–Њ 12 –Љ–Є–љ. –°–ї–µ–і—Г–µ—В –њ–Њ–Љ–љ–Є—В—М, —З—В–Њ –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Є–є —А–∞—Б—В–≤–Њ—А –і–Њ–ї–ґ–µ–љ –±—Л—В—М –Ї–Њ–Љ–љ–∞—В–љ–Њ–є —В–µ–Љ–њ–µ—А–∞—В—Г—А—Л, –Њ–Ї–Њ–ї–Њ 18-20¬∞.
5. –Т–Њ –≤—А–µ–Љ—П –≥–Є–њ–Њ—В–Њ–љ–Є—З–µ—Б–Ї–Њ–є –Њ–±—А–∞–±–Њ—В–Ї–Є –Ї–∞–љ–∞–ї—М—Ж—Л –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —А–∞–Ј –њ–µ—А–µ–і–≤–Є–≥–∞—О—В –њ–Є–љ—Ж–µ—В–Њ–Љ –Є–Ј –Њ–і–љ–Њ–≥–Њ —Г—З–∞—Б—В–Ї–∞ —З–∞—И–Ї–Є –Я–µ—В—А–Є –≤ –і—А—Г–≥–Њ–є, –∞ –Ј–∞—В–µ–Љ –њ–µ—А–µ–љ–Њ—Б—П—В –≤ —Д–Є–Ї—Б–∞—В–Њ—А, –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–є –Є–Ј —В—А—С—Е —З–∞—Б—В–µ–є —Н—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є –Њ–і–љ–Њ–є —З–∞—Б—В–Є –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л. –§–Є–Ї—Б–∞—Ж–Є—П –њ—А–Њ–≤–Њ–і–Є—В—Б—П –≤ —Б—В–µ–Ї–ї—П–љ–љ–Њ–Љ –±—О–Ї—Б–µ —Б –Ї—А—Л—И–Ї–Њ–є, –Њ–±—К–µ–Љ —Д–Є–Ї—Б–∞—В–Њ—А–∞ –Њ–Ї–Њ–ї–Њ 20 –Љ–ї.
6. –Ф–ї—П —Е–Њ—А–Њ—И–µ–є —Д–Є–Ї—Б–∞—Ж–Є–Є –≤–њ–Њ–ї–љ–µ –і–Њ—Б—В–∞—В–Њ—З–љ–Њ 15 –Љ–Є–љ., –њ–Њ—Б–ї–µ —З–µ–≥–Њ –Љ–Њ–ґ–љ–Њ –≥–Њ—В–Њ–≤–Є—В—М –њ—А–µ–њ–∞—А–∞—В—Л. –Ю–і–љ–∞–Ї–Њ –µ—Б–ї–Є –љ–µ—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –љ–µ–Љ–µ–і–ї–µ–љ–љ–Њ —Б–і–µ–ї–∞—В—М –њ—А–µ–њ–∞—А–∞—В—Л, —В–Њ –Ї–∞–љ–∞–ї—М—Ж—Л –Љ–Њ–ґ–љ–Њ —Б–Њ—Е—А–∞–љ–Є—В—М –≤ —Д–Є–Ї—Б–∞—В–Њ—А–µ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –њ–µ–і–µ–ї—М –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ –Њ—В 0 –і–Њ +4¬∞–°.
7. –Ф–ї—П –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Љ–Є–ї–ї–Є–ї–Є—В—А–Њ–≤ —Д–Є–Ї—Б–∞—В–Њ—А–∞ –≤–Љ–µ—Б—В–µ —Б –љ–µ–±–Њ–ї—М—И–Є–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ–Љ –Ї–∞–љ–∞–ї—М—Ж–µ–≤ –њ–Њ–Љ–µ—Й–∞—О—В –≤ –Љ–∞–ї–µ–љ—М–Ї—Г—О —З–∞—И–Ї—Г –Я–µ—В—А–Є. –Я—А–Є –њ–Њ–Љ–Њ—Й–Є –њ–∞—А—Л –њ—А–µ–њ–∞—А–Њ–≤–∞–ї—М–љ—Л—Е –Є–≥–ї –Ї–∞–љ–∞–ї—М—Ж—Л –Є–Ј–Љ–µ–ї—М—З–∞—О—В, –Њ—В–і–µ–ї—П—О—В –Є—Е –Њ—В —Д–Є–Ї—Б–∞—В–Њ—А–∞ –Є –њ–µ—А–µ–љ–Њ—Б—П—В –≤ 60%-–љ—Г—О —Г–Ї—Б—Г—Б–љ—Г—О –Ї–Є—Б–ї–Њ—В—Г, –љ–∞–ї–Є—В—Г—О –≤ –Љ–∞–ї–µ–љ—М–Ї—Г—О –∞–≥–≥–ї—О—В–Є–љ–∞—Ж–Є–Њ–љ–љ—Г—О –њ—А–Њ–±–Є—А–Ї—Г (–Њ–Ї–Њ–ї–Њ 0,5 –Љ–ї). –Ч–і–µ—Б—М –Ї–∞–љ–∞–ї—М—Ж—Л –±—Л—Б—В—А–Њ —Б—В–∞–љ–Њ–≤—П—В—Б—П –њ—А–Њ–Ј—А–∞—З–љ—Л–Љ–Є, —В–∞–Ї –Ї–∞–Ї —Б–њ–µ—А–Љ–∞—В–Њ–≥–Њ–љ–Є–∞–ї—М–љ—Л–µ –Ї–ї–µ—В–Ї–Є –≤—Л–њ–∞–і–∞—О—В –≤ —Б—Г—Б–њ–µ–љ–Ј–Є—О. –Э–µ–Ї–Њ—В–Њ—А–Њ–µ –њ–Њ—Б—В—Г–Ї–Є–≤–∞–љ–Є–µ –њ–Њ —Б—В–µ–љ–Ї–µ –њ—А–Њ–±–Є—А–Ї–Є –і–∞—С—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –њ–Њ–ї—Г—З–Є—В—М –Њ–і–љ–Њ—А–Њ–і–љ—Г—О –≤–Ј–≤–µ—Б—М.
8. –Ю–і–љ–Њ—А–Њ–і–љ—Г—О —Б—Г—Б–њ–µ–љ–Ј–Є—О –Ї–ї–µ—В–Њ–Ї –љ–∞–±–Є—А–∞—О—В –≤ –Љ–Є–Ї—А–Њ–њ–Є–њ–µ—В–Ї—Г –Є –Њ–і–љ—Г –Ї–∞–њ–ї—О –љ–∞–љ–Њ—Б—П—В –љ–∞ –Њ–±–µ–Ј–ґ–Є—А–µ–љ–љ–Њ–µ –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ, –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ –љ–∞–≥—А–µ—В–Њ–µ –і–Њ 60 ¬∞–°.. –Ґ–Њ—В—З–∞—Б –ґ–µ —Н—В—Г –Ї–∞–њ–ї—О –≤–љ–Њ–≤—М –≤–±–Є—А–∞—О—В –≤ –Љ–Є–Ї—А–Њ–њ–Є–њ–µ—В–Ї—Г –Є –њ–µ—А–µ–љ–Њ—Б—П—В –љ–∞ –і—А—Г–≥–Њ–µ –Љ–µ—Б—В–Њ —Н—В–Њ–≥–Њ –ґ–µ —Б—В–µ–Ї–ї–∞. –Ю–њ–µ—А–∞—Ж–Є—П –њ–Њ–≤—В–Њ—А—П–µ—В—Б—П 4-5 —А–∞–Ј –і–ї—П –Ї–∞–ґ–і–Њ–є –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ–Њ–є –Ї–∞–њ–ї–Є. –Я—А–Њ—Ж–µ—Б—Б –Љ–Њ–ґ–µ—В –і–ї–Є—В—М—Б—П –Є –і–Њ–ї—М—И–µ, –њ–Њ–Ї–∞ –љ—Г–ґ–љ–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Ї–ї–µ—В–Њ–Ї –љ–µ –±—Г–і–µ—В –љ–∞–љ–µ—Б–µ–љ–Њ –љ–∞ —Б—В–µ–Ї–ї–Њ. –Я–Њ—Б–ї–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е –Ї–∞–њ–µ–ї—М —Б—Г—Б–њ–µ–љ–Ј–Є—П –≤—Л–ї–Є–≤–∞–µ—В—Б—П, –љ–Њ–≤–Њ–µ —Б—В–µ–Ї–ї–Њ –Ј–∞—Б–µ–ї—П—О—В –Ї–ї–µ—В–Ї–∞–Љ–Є, –њ–Њ–ї—Г—З–µ–љ–љ—Л–Љ–Є –Є–Ј —Б–≤–µ–ґ–µ–є —В–Ї–∞–љ–Є. –Я—А–Є–љ—Ж–Є–њ, –љ–∞ –Ї–Њ—В–Њ—А–Њ–Љ –Њ—Б–љ–Њ–≤–∞–љ–Њ –њ–Њ–ї—Г—З–µ–љ–Є–µ –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤ —В–Њ–Љ, —З—В–Њ —Б—Г—Б–њ–µ–љ–Ј–Є—А–Њ–≤–∞–љ–љ—Л–µ –Ї–ї–µ—В–Ї–Є –Љ–Є–≥—А–Є—А—Г—О—В –Ї –њ–µ—А–Є—Д–µ—А–Є–Є —Б –Ї–∞–њ–ї–µ–є –Є –њ—А–Є –±—Л—Б—В—А–Њ–Љ –Њ—В—Б–∞—Б—Л–≤–∞–љ–Є–Є –Ї–∞–њ–ї–Є –Љ–Є–Ї—А–Њ–њ–Є–њ–µ—В–Ї–Њ–є –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –њ–Њ–і—Б—Л—Е–∞–љ–Є–µ —З–∞—Б—В–Є –Ї–ї–µ—В–Њ–Ї —В–Њ–љ–Ї–Є–Љ —Б–ї–Њ–µ–Љ, –∞ –љ–µ —Б–Ї—Г—З–µ–љ–љ–Њ, —З—В–Њ —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В —Е–Њ—А–Њ—И–µ–Љ—Г —А–∞–Ј–±—А–Њ—Б—Г —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –Ц–µ–ї–∞—В–µ–ї—М–љ–Њ –≥–Њ—В–Њ–≤–Є—В—М –Љ–∞–ї–µ–љ—М–Ї–Є–µ –њ–Њ—А—Ж–Є–Є –Љ–∞—В–µ—А–Є–∞–ї–∞, —В–∞–Ї –Ї–∞–Ї –њ–Њ—Б–ї–µ 5 –Љ–Є–љ. –њ—А–µ–±—Л–≤–∞–љ–Є—П –≤ 60%-–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В–µ –Ї–ї–µ—В–Ї–Є –љ–∞—З–Є–љ–∞—О—В —А–∞–Ј—А—Г—И–∞—В—М—Б—П. –Ш—Б–њ–Њ–ї—М–Ј—Г—П —Н—В–Њ—В –Љ–µ—В–Њ–і, –Љ–Њ–ґ–љ–Њ –љ–∞–±–ї—О–і–∞—В—М —А–∞–Ј–ї–Є—З–љ—Л–µ —Б—В–∞–і–Є–Є —Б–њ–µ—А–Љ–∞—В–Њ–≥–µ–љ–µ–Ј–∞. –Я—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ —Б—Е–Њ–і–љ–∞—П, –љ–Њ –±–Њ–ї–µ–µ —В—А—Г–і–Њ–µ–Љ–Ї–∞—П –Љ–µ—В–Њ–і–Є–Ї–∞ –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–Є—П –Ї–∞–Ї –Љ–µ–є–Њ—В–Є—З–µ—Б–Ї–Є—Е, —В–∞–Ї –Є –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Є—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Є–Ј —Б–µ–Љ–µ–љ–љ–Є–Ї–Њ–≤ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Њ–њ–Є—Б–∞–љ–∞ –Р.–Я. –Ф—Л–±–∞–љ–Њ–Љ (1970).
II
–†–Р–Ч–Ы–Ш–І–Э–Ђ–Х –°–Я–Ю–°–Ю–С–Ђ –Ю–Ъ–†–Р–°–Ъ–Ш –•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–• –Я–†–Х–Я–Р–†–Р–Ґ–Ю–Т
–Я–Њ–ї—Г—З–µ–љ–Є–µ —Е–Њ—А–Њ—И–Њ –Њ–Ї—А–∞—И–µ–љ–љ—Л—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —Б–Њ–±–Њ–є –Њ–і–Є–љ –Є–Ј –≤–∞–ґ–љ–µ–є—И–Є—Е —Н—В–∞–њ–Њ–≤ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞. –Я—А–Є –њ—А–∞–≤–Є–ї—М–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–µ –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ–Љ –Ї—А–∞—Б–Є—В–µ–ї–µ–є —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л —Б—В–∞–љ–Њ–≤—П—В—Б—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–≥–Њ –Ї—А–∞—Б–љ–Њ-—Д–Є–Њ–ї–µ—В–Њ–≤–Њ–≥–Њ —Ж–≤–µ—В–∞, –∞ –њ–Њ–і –Љ–Є–Ї—А–Њ—Б–Ї–Њ–њ–Њ–Љ —Б –Ј–µ–ї—С–љ—Л–Љ —Д–Є–ї—М—В—А–Њ–Љ, –Ї–Њ—В–Њ—А—Л–Љ —А–µ–Ї–Њ–Љ–µ–љ–і—Г–µ—В—Б—П –њ–Њ–ї—М–Ј–Њ–≤–∞—В—М—Б—П –њ—А–Є –њ—А–Њ—Б–Љ–Њ—В—А–µ –Є —Д–Њ—В–Њ–≥—А–∞—Д–Є—А–Њ–≤–∞–љ–Є–Є –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –≤—Л–≥–ї—П–і—П—В —З–µ—А–љ—Л–Љ–Є, —Б —З–µ—В–Ї–Є–Љ–Є –љ–µ—А–∞–Ј–Љ—Л—В—Л–Љ–Є –Ї—А–∞—П–Љ–Є –Є —Е–Њ—А–Њ—И–Њ —А–∞–Ј–ї–Є—З–Є–Љ—Л–Љ–Є —Е—А–Њ–Љ–∞—В–Є–і–∞–Љ–Є.
–Ю–Ъ–†–Р–°–Ъ–Р –Р–Ч–£–†-–≠–Ю–Ч–Ш–Э–Ю–Ь
–Т –Њ—В–і–µ–ї—М–љ—Л—Е —Д–ї–∞–Ї–Њ–љ–∞—Е –≥–Њ—В–Њ–≤—П—В—Б—П –Є—Б—Е–Њ–і–љ—Л–µ —А–∞—Б—В–≤–Њ—А—Л –Ї—А–∞—Б–Є—В–µ–ї–µ–є: 1) 100 –Љ–≥ —Б—Г—Е–Њ–≥–Њ –∞–Ј—Г—А–∞-II –љ–∞ 100 –Љ–ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л; 2) 100 –Љ–≥ —Б—Г—Е–Њ–≥–Њ –≤–Њ–і–Њ—А–∞—Б—В–≤–Њ—А–Є–Љ–Њ–≥–Њ —Н–Њ–Ј–Є–љ–∞ –љ–∞ 100 –Љ–ї –≤–Њ–і—Л. –Я—А–Є —Е—А–∞–љ–µ–љ–Є–Є —Н—В–Є—Е —А–∞—Б—В–≤–Њ—А–Њ–≤ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –Є—Е ¬Ђ–≤—Л–Ј—А–µ–≤–∞–љ–Є–µ¬ї, –њ–Њ—Н—В–Њ–Љ—Г –Ї—А–∞—Б–Ї–Є, –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–µ –Ј–∞—А–∞–љ–µ–µ, –Ї—А–∞—Б—П—В –ї—Г—З—И–µ, —З–µ–Љ —Б–≤–µ–ґ–Є–µ. –Э–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –њ–µ—А–µ–і –Њ–Ї—А–∞—Б–Ї–Њ–є –≥–Њ—В–Њ–≤–Є—В—Б—П —А–∞–±–Њ—З–Є–є —А–∞—Б—В–≤–Њ—А: —Б–Љ–µ—И–Є–≤–∞—О—В—Б—П 2 —З–∞—Б—В–Є –Є—Б—Е–Њ–і–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Н–Њ–Ј–Є–љ–∞, 3 —З–∞—Б—В–Є –Є—Б—Е–Њ–і–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –∞–Ј—Г—А–∞ –Є 5 —З–∞—Б—В–µ–є –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л. –Ф–ї—П –ї—Г—З—И–µ–≥–Њ –Њ–Ї—А–∞—И–Є–≤–∞–љ–Є—П –≤ –≥–Њ—В–Њ–≤—Л–є –Ї—А–∞—Б–Є—В–µ–ї—М –і–Њ–±–∞–≤–ї—П–µ—В—Б—П 0,5-1 –Љ–ї (–љ–∞ 100 –Љ–ї –Ї—А–∞—Б–Є—В–µ–ї—П) 0,1%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Г–≥–ї–µ–Ї–Є—Б–ї–Њ–≥–Њ –љ–∞—В—А–Є—П —В–∞–Ї, —З—В–Њ–±—Л —А–µ–∞–Ї—Ж–Є—П –Ї—А–∞—Б–Є—В–µ–ї—П –±—Л–ї–∞ –±–ї–Є–Ј–Ї–Њ–є –Ї –љ–µ–є—В—А–∞–ї—М–љ–Њ–є. –Я—А–µ–і–Љ–µ—В–љ—Л–µ —Б—В–µ–Ї–ї–∞ –њ–Њ–Љ–µ—Й–∞—О—В—Б—П –≤ —Б—В–∞–Ї–∞–љ—З–Є–Ї–Є —Б –Ї—А–∞—Б–Ї–Њ–є –Є –≤—Л–і–µ—А–ґ–Є–≤–∞—О—В—Б—П, –≤ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В –Ї–∞—З–µ—Б—В–≤–∞ –Ї—А–∞—Б–Є—В–µ–ї–µ–є, –Њ—В 5 –і–Њ 20 –Љ–Є–љ. –Я–Њ—Б–ї–µ —Н—В–Њ–≥–Њ —Б—В–µ–Ї–ї–∞ —Б–њ–Њ–ї–∞—Б–Ї–Є–≤–∞—О—В—Б—П –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ, –≤—Л—Б—Г—И–Є–≤–∞—О—В—Б—П –Є –њ—А–Њ–≤–Њ–і—П—В—Б—П —Б–љ–∞—З–∞–ї–∞ —З–µ—А–µ–Ј —Б–Љ–µ—Б—М –±—Г—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Є –Ї—Б–Є–ї–Њ–ї–∞ (1:2), –∞ –Ј–∞—В–µ–Љ —З–µ—А–µ–Ј —З–Є—Б—В—Л–є –Ї—Б–Є–ї–Њ–ї. –Я–Њ—Б–ї–µ —Н—В–Њ–≥–Њ –љ–∞ —Б—В–µ–Ї–ї–Њ –љ–∞–љ–Њ—Б–Є—В—Б—П –Ї–∞–њ–ї—П –Ї–∞–љ–∞–і—Б–Ї–Њ–≥–Њ –±–∞–ї—М–Ј–∞–Љ–∞ –Є –Њ–љ–Њ –Ј–∞–Ї—А—Л–≤–∞–µ—В—Б—П –њ–Њ–Ї—А–Њ–≤–љ—Л–Љ —Б—В–µ–Ї–ї–Њ–Љ.
–Я–Њ–Ї—А–Њ–≤–љ—Л–µ —Б—В–µ–Ї–ї–∞ –Ї–Є–њ—П—В—П—В –≤ –Љ—Л–ї—М–љ–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ, –Њ—В–Љ—Л–≤–∞—О—В –≥–Њ—А—П—З–µ–є –њ—А–Њ—В–Њ—З–љ–Њ–є, –∞ –Ј–∞—В–µ–Љ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–Њ–є –Є –Ї–ї–∞–і—Г—В –≤ —Б–Љ–µ—Б—М —Б–њ–Є—А—В–∞ —Б —Н—Д–Є—А–Њ–Љ –Є–ї–Є –≤ —З–Є—Б—В—Л–є —Б–њ–Є—А—В. –Ч–∞—В–µ–Љ –Ї–∞–ґ–і–Њ–µ —Б—В–µ–Ї–ї–Њ —В—Й–∞—В–µ–ї—М–љ–Њ –≤—Л—В–Є—А–∞–µ—В—Б—П —З–Є—Б—В–Њ–є, —Б–ї–Њ–ґ–µ–љ–љ–Њ–є –≤—З–µ—В–≤–µ—А–Њ –Љ–∞—А–ї–µ–є, –њ–Њ—Б–ї–µ —З–µ–≥–Њ –Є–Љ –Љ–Њ–ґ–љ–Њ –љ–∞–Ї—А—Л–≤–∞—В—М –њ—А–µ–њ–∞—А–∞—В.
–Ф–ї—П –Њ–Ї—А–∞—Б–Ї–Є –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Љ–Њ–ґ–љ–Њ –њ–Њ–ї—М–Ј–Њ–≤–∞—В—М—Б—П —В–∞–Ї–ґ–µ –≥–Њ—В–Њ–≤—Л–Љ –Ї—А–∞—Б–Є—В–µ–ї–µ–Љ –У–Є–Љ–Ј–∞ (–∞–Ј—Г—А-—Н–Њ–Ј–Є–љ –њ–Њ –†–Њ–Љ–∞–љ–Њ–≤—Б–Ї–Њ–Љ—Г). –Я—А–Є —Н—В–Њ–Љ –љ–∞ 100 –Љ–ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л –±–µ—А—С—В—Б—П 5 –Љ–ї –≥–Њ—В–Њ–≤–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ –У–Є–Љ–Ј–∞ –Є 2-3 –Љ–ї 0,1%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Г–≥–ї–µ–Ї–Є—Б–ї–Њ–≥–Њ –љ–∞—В—А–Є—П. –Я—А–µ–њ–∞—А–∞—В –њ–Њ–≥—А—Г–ґ–∞–µ—В—Б—П –≤ –Ї—А–∞—Б–Ї—Г –љ–∞ 5-10 –Љ–Є–љ., –Ј–∞—В–µ–Љ —Б–њ–Њ–ї–∞—Б–Ї–Є–≤–∞–µ—В—Б—П –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–Њ–є, –≤—Л—Б—Г—И–Є–≤–∞–µ—В—Б—П, –њ—А–Њ–≤–Њ–і–Є—В—Б—П —З–µ—А–µ–Ј —З–Є—Б—В—Л–є –Ї—Б–Є–ї–Њ–ї –Є –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤ –±–∞–ї—М–Ј–∞–Љ.
–Р–Ј—Г—А-—Н–Њ–Ј–Є–љ–Њ–Љ –Њ–і–Є–љ–∞–Ї–Њ–≤–Њ —Е–Њ—А–Њ—И–Њ –Њ–Ї—А–∞—И–Є–≤–∞—О—В—Б—П –Ї–∞–Ї –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Є–µ, —В–∞–Ї –Є –Љ–µ–є–Њ—В–Є—З–µ—Б–Ї–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л, –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–µ –Є–Ј –Ї–ї–µ—В–Њ—З–љ–Њ–є —Б—Г—Б–њ–µ–љ–Ј–Є–Є —Б –≤—Л–ґ–Є–≥–∞–љ–Є–µ–Љ —Д–Є–Ї—Б–∞—В–Њ—А–∞ –Є –±–µ–Ј –≤—Л–ґ–Є–≥–∞–љ–Є—П.
–Ю–Ъ–†–Р–°–Ъ–Р –Ю–†–°–Х–Ш–Э–Ю–Ь
–У–Њ—В–Њ–≤–Є—В—Б—П 2%-–љ—Л–є —А–∞—Б—В–≤–Њ—А –∞—Ж–µ—В–Њ—А—Б–µ–Є–љ–∞. –Ф–ї—П —Н—В–Њ–≥–Њ 2 –≥ —Б—Г—Е–Њ–≥–Њ –Њ—А—Б–µ–Є–љ–∞ —А–∞—Б—В–≤–Њ—А—П—О—В –≤ 45 –Љ–ї –≥–Њ—А—П—З–µ–є –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є –њ–Њ—Б–ї–µ –Њ—Е–ї–∞–ґ–і–µ–љ–Є—П –і–Њ–±–∞–≤–ї—П—О—В 55 –Љ–ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л. –Ь–Њ–ґ–љ–Њ –≥–Њ—В–Њ–≤–Є—В—М –Њ—А—Б–µ–Є–љ –љ–µ –љ–∞ 45%-–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є, –∞ –љ–∞ 60%-–љ–Њ–є –њ—А–Њ–њ–Є–Њ–љ–Њ–≤–Њ–є –Є–ї–Є –Љ–Њ–ї–Њ—З–љ–Њ–є –Ї–Є—Б–ї–Њ—В–µ, –Є–ї–Є –љ–∞ —Б–Љ–µ—Б—П—Е –Љ–Њ–ї–Њ—З–љ–Њ–є –Є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В (–ї–∞–Ї—В–∞—Ж–µ—В-–Њ—А—Б–µ–Є–љ). –Т –њ–Њ—Б–ї–µ–і–љ–µ–Љ —Б–ї—Г—З–∞–µ —Б–Љ–µ—И–Є–≤–∞—О—В—Б—П —А–∞–≤–љ—Л–µ —З–∞—Б—В–Є 60% -–љ–Њ–є –Љ–Њ–ї–Њ—З–љ–Њ–є –Є –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В, –њ–Њ–і–Њ–≥—А–µ–≤–∞—О—В—Б—П –Є –≤ —Н—В–Њ–є –ґ–Є–і–Ї–Њ—Б—В–Є —А–∞—Б—В–≤–Њ—А—П–µ—В—Б—П –Њ—А—Б–µ–Є–љ (–Є–Ј —А–∞—Б—З–µ—В–∞ 2 –≥ –љ–∞ 100 –Љ–ї —Б–Љ–µ—Б–Є –Ї–Є—Б–ї–Њ—В). –Ч–∞—В–µ–Љ —Б–≤–µ–ґ–µ–њ—А–Њ—Д–Є–ї—М—В—А–Њ–≤–∞–љ–љ—Л–є —А–∞—Б—В–≤–Њ—А –Њ—А—Б–µ–Є–љ–∞ –Ї–∞–њ–∞—О—В –љ–∞ –њ—А–µ–і–Љ–µ—В–љ–Њ–µ —Б—В–µ–Ї–ї–Њ –Є –њ–Њ–Ї—А—Л–≤–∞—О—В –њ–Њ–Ї—А–Њ–≤–љ—Л–Љ —Б—В–µ–Ї–ї–Њ–Љ. –Ш–Ј–ї–Є—И–µ–Ї –Њ—А—Б–µ–Є–љ–∞ —Г–і–∞–ї—П—О—В —Д–Є–ї—М—В—А–Њ–≤–∞–ї—М–љ–Њ–є –±—Г–Љ–∞–≥–Њ–є –Є –њ–Њ –Ї—А–∞—О –њ–Њ–Ї—А–Њ–≤–љ–Њ–≥–Њ —Б—В–µ–Ї–ї–∞ –Ј–∞–Ї–ї–µ–Є–≤–∞—О—В –µ–≥–Њ –≥–Њ—А—П—З–Є–Љ –њ–∞—А–∞—Д–Є–љ–Њ–Љ (–Є–ї–Є —Б–Љ–µ—Б—М—О –≤–Њ—Б–Ї–∞ —Б –њ–∞—А–∞—Д–Є–љ–Њ–Љ). –Т —В–∞–Ї–Њ–Љ –≤–Є–і–µ —Б—В–µ–Ї–ї–∞ –њ–Њ–Љ–µ—Й–∞—О—В—Б—П –≤ –Ї–∞–Љ–µ—А—Г —Б –њ–∞—А–∞–Љ–Є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л (–љ–∞–њ—А–Є–Љ–µ—А, –≤ —З–∞—И–Ї—Г –Я–µ—В—А–Є, –љ–∞ –і–љ–Њ –Ї–Њ—В–Њ—А–Њ–є –њ–Њ–ї–Њ–ґ–µ–љ–∞ —Д–Є–ї—М—В—А–Њ–≤–∞–ї—М–љ–∞—П –±—Г–Љ–∞–≥–∞, —Б–Љ–Њ—З–µ–љ–љ–∞—П —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є) –Є —Б—В–∞–≤—П—В—Б—П –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї –љ–∞ 2-3 —Б—Г—В–Њ–Ї. –Ч–∞—В–µ–Љ –њ–Њ–Ї—А–Њ–≤–љ–Њ–µ —Б—В–µ–Ї–ї–Њ –≤–Љ–µ—Б—В–µ —Б –њ–∞—А–∞—Д–Є–љ–Њ–Љ –Њ—Б—В–Њ—А–Њ–ґ–љ–Њ —Г–і–∞–ї—П–µ—В—Б—П –±—А–Є—В–≤–Њ–є, –Є –њ—А–µ–њ–∞—А–∞—В—Л –њ—А–Њ–≤–Њ–і—П—В—Б—П –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ —З–µ—А–µ–Ј 45%-–љ—Г—О —Г–Ї—Б—Г—Б–љ—Г—О –Ї–Є—Б–ї–Њ—В—Г, –±—Г—В–Є–ї–Њ–≤—Л–є —Б–њ–Є—А—В –Є –і–≤–∞ –Ї—Б–Є–ї–Њ–ї–∞, –њ–Њ—Б–ї–µ —З–µ–≥–Њ –Ј–∞–Ї–ї—О—З–∞—О—В—Б—П –≤ –±–∞–ї—М–Ј–∞–Љ.
–•—А–∞–љ–Є—В—М —А–∞—Б—В–≤–Њ—А –Њ—А—Б–µ–Є–љ–∞ —Б–ї–µ–і—Г–µ—В –≤ —В–µ–Љ–љ–Њ–є –±—Г—В—Л–ї–Ї–µ —Б –њ—А–Є—В–µ—А—В–Њ–є –њ—А–Њ–±–Ї–Њ–є, –ґ–µ–ї–∞—В–µ–ї—М–љ–Њ –љ–∞ —Е–Њ–ї–Њ–і—Г (–њ—А–Є 4¬∞–° –Љ–Њ–ґ–љ–Њ —Е—А–∞–љ–Є—В—М –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Љ–µ—Б—П—Ж–µ–≤), –∞ —Д–Є–ї—М—В—А–Њ–≤–∞—В—М —Б–ї–µ–і—Г–µ—В –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –њ–µ—А–µ–і —Г–њ–Њ—В—А–µ–±–ї–µ–љ–Є–µ–Љ. –Ы—Г—З—И–µ –≤—Б–µ–≥–Њ –Њ–Ї—А–∞—И–Є–≤–∞–µ—В —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л –Њ—А—Б–µ–Є–љ —Д–Є—А–Љ—Л –Ь–µ—А–Ї (Merck, Darmstatd, Orcein 7091) –Є–ї–Є —Д–Є—А–Љ—Л –У–µ—А—А (–Ґ. Gurr, London). –Я—А–µ–њ–∞—А–∞—В—Л, –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–µ —Б –≤—Л–ґ–Є–≥–∞–љ–Є–µ–Љ —Д–Є–Ї—Б–∞—В–Њ—А–∞, –Њ–Ї—А–∞—И–Є–≤–∞—О—В—Б—П –Њ—А—Б–µ–Є–љ–Њ–Љ –њ–ї–Њ—Е–Њ.
–Ю–Ъ–†–Р–°–Ъ–Р –Я–Ю –§–Б–Ы–ђ–У–Х–Э–£
1 –≥ —Д—Г–Ї—Б–Є–љ–∞ –Ї–Є–њ—П—В—П—В –≤ —В–µ—З–µ–љ–Є–µ 5 –Љ–Є–љ. –≤ 200 –Љ–ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л, –Ј–∞—В–µ–Љ –Њ—Е–ї–∞–ґ–і–∞—О—В –≤ –њ—А–Њ—В–Њ—З–љ–Њ–є –≤–Њ–і–µ –њ–Њ–і –Ї—А–∞–љ–Њ–Љ –Є —Д–Є–ї—М—В—А—Г—О—В —З–µ—А–µ–Ј –±—Г–Љ–∞–ґ–љ—Л–є —Д–Є–ї—М—В—А. –Ъ —Д–Є–ї—М—В—А–∞—В—Г –і–Њ–±–∞–≤–ї—П—О—В 20 –Љ–ї 1%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є 1 –≥ –±–Є—Б—Г–ї—М—Д–Є—В–∞ –љ–∞—В—А–Є—П. –Ъ—А–∞—Б–Ї—Г —Е—А–∞–љ—П—В –≤ —Е–Њ–ї–Њ–і–Є–ї—М–љ–Є–Ї–µ –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ +(2-3)¬∞–°. –Я—А–µ–њ–∞—А–∞—В—Л –Њ–Ї—А–∞—И–Є–≤–∞—О—В –≤ —В–µ—З–µ–љ–Є–µ 1 —З–∞—Б–∞. –Я–µ—А–µ–і –Њ–Ї—А–∞—Б–Ї–Њ–є –њ—А–µ–њ–∞—А–∞—В—Л –і–Њ–ї–ґ–љ—Л –±—Л—В—М –њ–Њ–і–≤–µ—А–≥–љ—Г—В—Л –≥–Є–і—А–Њ–ї–Є–Ј—Г –≤ 1%-–љ–Њ–Љ —А–∞—Б—В–≤–Њ—А–µ —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л –≤ —В–µ—З–µ–љ–Є–µ 12 –Љ–Є–љ. –≤ —В–µ—А–Љ–Њ—Б—В–∞—В–µ –њ—А–Є 58¬∞–°. –Я–Њ—Б–ї–µ –≥–Є–і—А–Њ–ї–Є–Ј–∞ –њ—А–µ–њ–∞—А–∞—В—Л –њ—А–Њ–Љ—Л–≤–∞—О—В 2 —А–∞–Ј–∞ –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ –Є –њ—А–Є—Б—В—Г–њ–∞—О—В –Ї –Њ–Ї—А–∞—Б–Ї–µ.
–Ю–Ъ–†–Р–°–Ъ–Р –У–Х–Ь–Р–Ґ–Ю–Ъ–°–Ш–Ы–Ш–Э–Ю–Ь –Я–Ю –≠–†–Ы–Ш–•–£
–°–Љ–µ—И–∞—В—М: 2 –≥ –≥–µ–Љ–∞—В–Њ–Ї—Б–Є–ї–Є–љ–∞ –≠—А–ї–Є—Е–∞, 100 –Љ–ї 95%-–љ–Њ–≥–Њ —Н—В–Є–ї–Њ–≤–Њ–≥–Њ —Б–њ–Є—А—В–∞, 100 –Љ–ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л, 100 –Љ–ї –≥–ї–Є—Ж–µ—А–Є–љ–∞, 3 –≥ –Ї–∞–ї–Є–є–љ—Л—Е –Ї–≤–∞—Б—Ж–Њ–≤ –Є 10 –Љ–ї –ї–µ–і—П–љ–Њ–є —Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л. –°–Љ–µ—Б—М –і–µ—А–ґ–∞—В—М –і–≤–µ –љ–µ–і–µ–ї–Є –љ–∞ —Б–≤–µ—В—Г.
–Я–Њ—Б–ї–µ –≥–Є–і—А–Њ–ї–Є–Ј–∞ (—Б–Љ. –Њ–Ї—А–∞—Б–Ї—Г –њ–Њ –§—С–ї—М–≥–µ–љ—Г), –њ—А–Њ–Љ—Л–≤–Ї–Є –њ–Њ–і –Ї—А–∞–љ–Њ–Љ –Є –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ –њ—А–µ–њ–∞—А–∞—В –Њ–њ—Г—Б–Ї–∞—О—В –≤ –Ї—А–∞—Б–Ї—Г –Є –і–µ—А–ґ–∞—В –Њ—В 10 –і–Њ 30 –Љ–Є–љ. –Х—Б–ї–Є –≥–µ–Љ–∞—В–Њ–Ї—Б–Є–ї–Є–љ —Б—В–∞—А—Л–є, —В–Њ –њ—А–µ–њ–∞—А–∞—В—Л –Љ–Њ–ґ–љ–Њ –Ї—А–∞—Б–Є—В—М –≤ —В–µ—З–µ–љ–Є–µ 10 –Љ–Є–љ. –Ф–∞–ї–µ–µ —Б–ї–µ–і—Г–µ—В –њ—А–Њ–Љ—Л–≤–Ї–∞ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–Њ–є –Є –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–Є–µ –≤ —Б–њ–Є—А—В–µ —Б —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є –≤ —В–µ—З–µ–љ–Є–µ 1-2 –Љ–Є–љ. (–љ–∞ 100 –Љ–ї 70%-–љ–Њ–≥–Њ —Б–њ–Є—А—В–∞ –Њ–Ї–Њ–ї–Њ 1 –Љ–ї 1%-–љ–Њ–≥–Њ —А–∞—Б—В–≤–Њ—А–∞ —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л). –Я–Њ—Б–ї–µ —Н—В–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В—Л –њ—А–Њ–Љ—Л–≤–∞—О—В –≤ –њ—А–Њ—В–Њ—З–љ–Њ–є –≤–Њ–і–µ 20-30 –Љ–Є–љ. –Х—Б–ї–Є –Њ–Ї—А–∞—И–Є–≤–∞—О—В –і–∞–≤–ї–µ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л —Е—А–Њ–Љ–Њ—Б–Њ–Љ, —В–Њ –њ–Њ—Б–ї–µ –њ—А–Њ–Љ—Л–≤–Ї–Є —Б–ї–µ–і—Г–µ—В –њ—А–Њ-–≤–Њ–і–Ї–∞ —З–µ—А–µ–Ј —Б–њ–Є—А—В—Л –Є –Ї—Б–Є–ї–Њ–ї:
1. 96%-–љ—Л–є —Б–њ–Є—А—В (1) вАУ 1 –Љ–Є–љ.
2. 96%-–љ—Л–є —Б–њ–Є—А—В (2) вАУ 1 –Љ–Є–љ.
3. –Р–±—Б–Њ–ї—О—В–љ—Л–є —Б–њ–Є—А—В (1) вАУ 1 –Љ–Є–љ.
4. –Р–±—Б–Њ–ї—О—В–љ—Л–є —Б–њ–Є—А—В (2) вАУ 1 –Љ–Є–љ.
5. –Ъ—Б–Є–ї–Њ–ї (1) вАУ 1 –Љ–Є–љ.
6. –Ъ—Б–Є–ї–Њ–ї (2) вАУ1 –Љ–Є–љ. (–Љ–Њ–ґ–љ–Њ –і–µ—А–ґ–∞—В—М –Є 2-3 –і–љ—П).
–Я–Њ—Б–ї–µ –њ—А–Њ–≤–Њ–і–Ї–Є –љ–∞ –њ—А–µ–њ–∞—А–∞—В –љ–∞–љ–Њ—Б–Є—В—Б—П –Ї–∞–њ–ї—П –Ї–∞–љ–∞–і—Б–Ї–Њ–≥–Њ –±–∞–ї—М–Ј–∞–Љ–∞ –Є –Ј–∞–Ї—А—Л–≤–∞–µ—В—Б—П —З–Є—Б—В—Л–Љ —В–Њ–љ–Ї–Є–Љ –њ–Њ–Ї—А–Њ–≤–љ—Л–Љ —Б—В–µ–Ї–ї–Њ–Љ.
–Ф–Ш–§–§–Х–†–Х–Э–¶–Ш–Р–Ы–ђ–Э–Р–ѓ –Ю–Ъ–†–Р–°–Ъ–Р –•–†–Ю–Ь–Ю–°–Ю–Ь
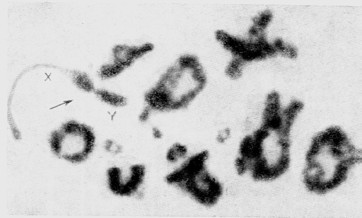
–Ь–µ—В–Њ–і—Л –і–Є—Д—Д–µ—А–µ–љ—Ж–Є–∞–ї—М–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ —З–µ–ї–Њ–≤–µ–Ї–∞ –Є –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, —И–Є—А–Њ–Ї–Њ —А–∞–Ј—А–∞–±–∞—В—Л–≤–∞–µ–Љ—Л–µ –≤ –њ–Њ—Б–ї–µ–і–љ–Є–µ –≥–Њ–і—Л —Ж–Є—В–Њ–ї–Њ–≥–∞–Љ–Є –≤—Б–µ–≥–Њ –Љ–Є—А–∞, –Њ—В–Ї—А—Л–≤–∞—О—В –њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ –љ–Њ–≤—Л–µ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –≤ —Ж–Є—В–Њ–≥–µ–љ–µ—В–Є–Ї–µ –ґ–Є–≤–Њ—В–љ—Л—Е. –°—Г—В—М —Н—В–Є—Е –Љ–µ—В–Њ–і–Њ–≤ –≤ —В–Њ–Љ, —З—В–Њ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л, –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ –Њ–±—А–∞–±–Њ—В–∞–љ–љ—Л–µ —А–∞–Ј–ї–Є—З–љ—Л–Љ–Є –≤–µ—Й–µ—Б—В–≤–∞–Љ–Є, –љ–∞—З–Є–љ–∞—О—В —Б–≤—П–Ј—Л–≤–∞—В—М –Ї—А–∞—Б–Є—В–µ–ї–Є (—Д–ї—Г–Њ—А–µ—Б—Ж–µ–љ—В–љ—Л–µ –Є–ї–Є –Ї—А–∞—Б–Є—В–µ–ї—М –У–Є–Љ–Ј–∞) –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ–Њ –њ–Њ –і–ї–Є–љ–µ –Ї–∞–ґ–і–Њ–є —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л. –Ю–Ї—А–∞—И–µ–љ–љ—Л–µ —В–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л –≤—Л–≥–ї—П–і—П—В –Ї–∞–Ї –±—Л ¬Ђ–њ–Њ–ї–Њ—Б–∞—В—Л–Љ–Є¬ї (banding), –њ—А–Є—З—С–Љ —Н—В–Є –њ–Њ–ї–Њ—Б—Л —А–∞–Ј–ї–Є—З–љ—Л –њ–Њ —И–Є—А–Є–љ–µ –Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є, –∞ —Е–∞—А–∞–Ї—В–µ—А –њ–Њ–њ–µ—А–µ—З–љ–Њ–є –Є—Б—З–µ—А—З–µ–љ–љ–Њ—Б—В–Є –Њ–Ї–∞–Ј—Л–≤–∞–µ—В—Б—П —Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Є–Љ –і–ї—П –Ї–∞–ґ–і–Њ–є –њ–∞—А—Л —Е—А–Њ–Љ–Њ—Б–Њ–Љ (—Б–Љ. —А–Є—Б.3).
–Я–Њ —Е–∞—А–∞–Ї—В–µ—А—Г –њ–Њ–ї—Г—З–∞–µ–Љ–Њ–є –Ї–∞—А—В–Є–љ—Л –Є—Б—З–µ—А—З–µ–љ–љ–Њ—Б—В–Є –Є –њ–Њ —В–Є–њ—Г –Њ–±—А–∞–±–Њ—В–Ї–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Љ–µ—В–Њ–і—Л –і–Є—Д—Д–µ—А–µ–љ—Ж–Є–∞–ї—М–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є –њ—А–Є–љ—П—В–Њ —Г—Б–ї–Њ–≤–љ–Њ —А–∞–Ј–і–µ–ї—П—В—М –љ–∞ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –≥—А—Г–њ–њ:
1) –Њ–Ї—А–∞—Б–Ї–∞ —Б –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ–Љ —Д–ї—Г–Њ—А–µ—Б—Ж–µ–љ—В–љ—Л—Е –Ї—А–∞—Б–Є—В–µ–ї–µ–є, –≤ –Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ–Њ–є —В–µ—А–Љ–Є–љ–Њ–ї–Њ–≥–Є–Є вАУ Q-bands (Q-–њ–Њ–ї–Њ—Б—Л) (Zech, 1969; Casperson et al., 1970; Lin et al, 1971, –Є –і—А.);
2) –Њ–Ї—А–∞—Б–Ї–∞, –≤—Л—П–≤–ї—П—О—Й–∞—П –Ї–Њ–љ—Б—В–Є—В—Г—В–Є–≤–љ—Л–є –≥–µ—В–µ—А–Њ—Е—А–Њ–Љ–∞—В–Є–љ вАУ C-bands (–°-–њ–Њ–ї–Њ—Б—Л) (Pardue, G–∞ll, 1970; Arrighi, Hsu, 1971, –Є –і—А.);
3) –Њ–Ї—А–∞—Б–Ї–∞ –Ї—А–∞—Б–Є—В–µ–ї–µ–Љ –У–Є–Љ–Ј–∞, —Б —А–∞–Ј–ї–Є—З–љ—Л–Љ–Є –њ—А–µ–і–Њ–±—А–∞–±–Њ—В–Ї–∞–Љ–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ вАУ G-bands (G-–њ–Њ–ї–Њ—Б—Л) (Evans et al., 1971; Seabright, 1971; Schnedl, 1971, –Є –і—А.).
–І–Є—Б–ї–Њ –Љ–µ—В–Њ–і–Є–Ї –Ї–∞–ґ–і–Њ–≥–Њ –Ї–ї–∞—Б—Б–∞ –≤–µ–ї–Є–Ї–Њ –Є –љ–µ–њ—А–µ—А—Л–≤–љ–Њ —А–∞—Б—В—С—В.
–°–ї–µ–і—Г–µ—В –Ј–∞–Љ–µ—В–Є—В—М, —З—В–Њ —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–Є–µ —Б–≤–µ—В—П—Й–Є—Е—Б—П –і–Є—Б–Ї–Њ–≤ –≤ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞—Е, –Њ–Ї—А–∞—И–µ–љ–љ—Л—Е —Д–ї—Г–Њ—А–µ—Б—Ж–µ–љ—В–љ—Л–Љ–Є –Ї—А–∞—Б–Є—В–µ–ї—П–Љ–Є, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, —Б–Њ–≤–њ–∞–і–∞–µ—В —Б —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–Є–µ–Љ —В–µ–Љ–љ—Л—Е –і–Є—Б–Ї–Њ–≤, –њ–Њ–ї—Г—З–∞–µ–Љ—Л—Е –њ—А–Є –Њ–Ї—А–∞—Б–Ї–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Ї—А–∞—Б–Є—В–µ–ї–µ–Љ –У–Є–Љ–Ј–∞, —Б —В–Њ–є —А–∞–Ј–љ–Є—Ж–µ–є, —З—В–Њ —Д–ї—Г–Њ—А–µ—Б—Ж–µ–љ—В–љ—Л–µ –Ї—А–∞—Б–Є—В–µ–ї–Є –≤—Л—П–≤–ї—П—О—В –±–Њ–ї–µ–µ –≥—А—Г–±—Л–є —А–Є—Б—Г–љ–Њ–Ї –Є—Б—З–µ—А—З–µ–љ–љ–Њ—Б—В–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –°–Њ–Ј–і–∞—С—В—Б—П —В–∞–Ї–Њ–µ –≤–њ–µ—З–∞—В–ї–µ–љ–Є–µ, —З—В–Њ —А–∞–Ј–ї–Є—З–љ—Л–µ –Љ–µ—В–Њ–і—Л –њ–Њ–ї—Г—З–µ–љ–Є—П G-–њ–Њ–ї–Њ—Б (–Њ–Ї—А–∞—Б–Ї–Њ–є –њ–Њ –У–Є–Љ–Ј–∞) –і–∞—О—В –µ—Б–ї–Є –љ–µ —Б–Њ–≤–µ—А—И–µ–љ–љ–Њ —Б—Е–Њ–і–љ—Л–µ, —В–Њ –≤–њ–Њ–ї–љ–µ —Б–Њ–њ–Њ—Б—В–∞–≤–Є–Љ—Л–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л.
–Ю–Ї—А–∞—Б–Ї–∞, –≤—Л—П–≤–ї—П—О—Й–∞—П –Ї–Њ–љ—Б—В–Є—В—Г—В–Є–≤–љ—Л–є –≥–µ—В–µ—А–Њ—Е—А–Њ–Љ–∞—В–Є–љ (–°-–њ–Њ–ї–Њ—Б—Л), –≤ –±–Њ–ї—М—И–Є–љ—Б—В–≤–µ —Б–ї—Г—З–∞–µ–≤ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–µ—В –≥–µ—В–µ—А–Њ—Е—А–Њ–Љ–∞—В–Є–љ–Њ–≤—Л–µ –±–ї–Њ–Ї–Є, –ї–Њ–Ї–∞–ї–Є–Ј–Њ–≤–∞–љ–љ—Л–µ –≤ —Ж–µ–љ—В—А–Њ–Љ–µ—А–љ—Л—Е –Є —В–µ–ї–Њ–Љ–µ—А–љ—Л—Е —А–∞–є–Њ–љ–∞—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ, –љ–Њ –Є–љ–Њ–≥–і–∞ –≤—Л—П–≤–ї—П—О—В—Б—П –≥–µ—В–µ—А–Њ—Е—А–Њ–Љ–∞—В–Є–љ–Њ–≤—Л–µ –Њ–±–ї–∞—Б—В–Є –≤ —А–∞–Ј–ї–Є—З–љ—Л—Е —Г—З–∞—Б—В–Ї–∞—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ–ї–µ—З –Є–ї–Є –і–∞–ґ–µ –њ–Њ–ї–љ–Њ—Б—В—М—О –≥–µ—В–µ—А–Њ—Е—А–Њ–Љ–∞—В–Є—З–µ—Б–Ї–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л.
–Ь–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л, –≤—Л–Ј—Л–≤–∞—О—Й–Є–µ –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є–µ —Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Њ–є –њ–Њ–њ–µ—А–µ—З–љ–Њ–є –Є—Б—З–µ—А—З–µ–љ–љ–Њ—Б—В–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ, –µ—Й—С —Б–Њ–≤–µ—А—И–µ–љ–љ–Њ –љ–µ —П—Б–љ—Л. –Я—А–µ–і–њ–Њ–ї–∞–≥–∞–µ—В—Б—П –ї–Є—И—М, —З—В–Њ —Н—В–Є —П–≤–ї–µ–љ–Є—П —Б–≤—П–Ј–∞–љ—Л —Б –њ—А–Њ—Ж–µ—Б—Б–∞–Љ–Є –і–µ–љ–∞—В—Г—А–∞—Ж–Є–Є –±–µ–ї–Ї–Њ–≤—Л—Е –Є–ї–Є –Ф–Э–Ъ-–Њ–≤—Л—Е —Б—В—А—Г–Ї—В—Г—А —Е—А–Њ–Љ–Њ—Б–Њ–Љ –њ—А–Є —А–∞–Ј–ї–Є—З–љ—Л—Е –Њ–±—А–∞–±–Њ—В–Ї–∞—Е (—Б–Њ–ї—П–Љ–Є, —В—А–Є–њ—Б–Є–љ–Њ–Љ, —А–∞—Б—В–≤–Њ—А–∞–Љ–Є –Љ–Њ—З–µ–≤–Є–љ—Л –Є —В. –і.) –Є –Њ–±—А–∞—В–љ–Њ–є —З–∞—Б—В–Є—З–љ–Њ–є –Є—Е –і–µ–љ–∞—В—Г—А–∞—Ж–Є–Є. –Э–µ–Ї–Њ—В–Њ—А—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–Є —Б–≤—П–Ј—Л–≤–∞—О—В –і–Є—Д—Д–µ—А–µ–љ—Ж–Є–∞–ї—М–љ—Г—О –Њ–Ї—А–∞—Б–Ї—Г —Б –љ–µ—А–∞–≤–љ–Њ–Љ–µ—А–љ—Л–Љ —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–Є–µ–Љ –њ–Њ –і–ї–Є–љ–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ —Г—З–∞—Б—В–Ї–Њ–≤ —А–µ–њ–µ—В–Є—А–Њ–≤–∞–љ–љ–Њ–є –Ф–Э–Ъ (—В. –µ. –Ф–Э–Ъ —Б –њ–Њ–≤—В–Њ—А—П—О—Й–Є–Љ–Є—Б—П –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В—П–Љ–Є). –Ю–і–љ–∞–Ї–Њ –њ–Њ–Ї–∞ –≤—Б–µ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ –≥–Є–њ–Њ—В–µ–Ј—Л —В–∞–Ї–Њ–≥–Њ —А–Њ–і–∞ –Њ—Б—В–∞—О—В—Б—П –≤ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–є —Б—В–µ–њ–µ–љ–Є —Б–њ–µ–Ї—Г–ї—П—В–Є–≤–љ—Л–Љ–Є.
–Ґ–µ–Љ –љ–µ –Љ–µ–љ–µ–µ, —В–Њ—В —Д–∞–Ї—В, —З—В–Њ —Е–∞—А–∞–Ї—В–µ—А –њ–Њ–њ–µ—А–µ—З–љ–Њ–є –Є—Б—З–µ—А—З–µ–љ–љ–Њ—Б—В–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ —Б–њ–µ—Ж–Є—Д–Є—З–µ–љ –і–ї—П –Ї–∞–ґ–і–Њ–є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–є –њ–∞—А—Л, –њ–Њ—З—В–Є –љ–µ –Ј–∞–≤–Є—Б–Є—В –Њ—В —Б—В–µ–њ–µ–љ–Є —Б–њ–Є—А–∞–ї–Є–Ј–∞—Ж–Є–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ (–≤ —Б–Є–ї—М–љ–Њ —Б–њ–Є—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ—Л—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞—Е –љ–µ—А–∞–Ј–ї–Є—З–Є–Љ—Л –љ–µ–Ї–Њ—В–Њ—А—Л–µ —В–Њ–љ–Ї–Є–µ –і–Є—Б–Ї–Є), –∞ —В–∞–Ї–ґ–µ —Б–Њ—Е—А–∞–љ—П–µ—В—Б—П –≤ –њ–µ—А–µ—Б—В—А–Њ–µ–љ–љ—Л—Е —Г—З–∞—Б—В–Ї–∞—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ, –њ–Њ–Ј–≤–Њ–ї—П–µ—В, –≤–Њ-–њ–µ—А–≤—Л—Е, —Б—В—А–Њ–≥–Њ –Є–і–µ–љ—В–Є—Д–Є—Ж–Є—А–Њ–≤–∞—В—М –≤—Б–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ–∞—А—Л –≤ –Ї–∞—А–Є–Њ—В–Є–њ–µ –Є, –≤–Њ-–≤—В–Њ—А—Л—Е, –њ—А–Њ—Б–ї–µ–і–Є—В—М —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –њ–µ—А–µ—Б—В—А–Њ–є–Ї–Є, —Б—А–∞–≤–љ–Є–≤–∞—П –њ—А–Є —Н—В–Њ–Љ –Ї–∞–Ї –Ї–∞—А–Є–Њ—В–Є–њ—Л –Њ–і–љ–Њ–≥–Њ –≤–Є–і–∞, —В–∞–Ї –Є —А–∞–Ј–љ—Л—Е –≤–Є–і–Њ–≤ —А–Њ–і—Б—В–≤–µ–љ–љ—Л—Е —Д–Њ—А–Љ –ґ–Є–≤–Њ—В–љ—Л—Е –Є –≤—Л—П–≤–ї—П—П –≥–Њ–Љ–Њ–ї–Њ–≥–Є—З–љ—Л–µ –Є –њ–µ—А–µ—Б—В—А–Њ–µ–љ–љ—Л–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л. –Т—Б–µ —Н—В–Њ –Њ—В–Ї—А—Л–≤–∞–µ—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –±–Њ–ї–µ–µ –≥–ї—Г–±–Њ–Ї–Њ–≥–Њ –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б —В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л–Љ–Є –Љ–µ—В–Њ–і–∞–Љ–Є –Ї–∞—А–Є–Њ—В–∞–Ї—Б–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –Є–Ј—Г—З–µ–љ–Є—П —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ—Л—Е —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–є –Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є, –≤ —В–µ—Е —Б–ї—Г—З–∞—П—Е, –Ї–Њ–≥–і–∞ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ—Л–є –≥–Є–±—А–Є–і–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–є –∞–љ–∞–ї–Є–Ј –љ–µ–≤–Њ–Ј–Љ–Њ–ґ–µ–љ –Є–Ј-–Ј–∞ —Б—А–∞–≤–љ–Є—В–µ–ї—М–љ–Њ –Њ—В–і–∞–ї—С–љ–љ–Њ–≥–Њ —Д–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ —А–Њ–і—Б—В–≤–∞ —Б–Њ–њ–Њ—Б—В–∞–≤–ї—П–µ–Љ—Л—Е —Д–Њ—А–Љ.
–Я–Њ—Б–Ї–Њ–ї—М–Ї—Г –Љ–µ—В–Њ–і—Л –Њ–Ї—А–∞—Б–Ї–Є –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ –Њ–±—А–∞–±–Њ—В–∞–љ–љ—Л—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Ї—А–∞—Б–Є—В–µ–ї–µ–Љ –У–Є–Љ–Ј–∞ –њ–Њ–Ј–≤–Њ–ї—П—О—В –≤—Л—П–≤–ї—П—В—М –љ–∞–Є–±–Њ–ї–µ–µ —В–Њ–љ–Ї—Г—О –Ї–∞—А—В–Є–љ—Г —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–є –Є—Б—З–µ—А—З–µ–љ–љ–Њ—Б—В–Є –Є –і–∞—О—В –ї—Г—З—И–Є–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л, –њ—А–Є–≤–µ–і—С–Љ –Њ–і–љ—Г –Є–Ј —Н—В–Є—Е –Љ–µ—В–Њ–і–Є–Ї, —А–∞–Ј—А–∞–±–Њ—В–∞–љ–љ—Г—О –°.–Ш. –†–∞–і–ґ–∞–±–ї–Є (–†–∞–і–ґ–∞–±–ї–Є, –Ъ—А—О–Ї–Њ–≤–∞, 1973).
–¶–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –њ—А–µ–њ–∞—А–∞—В—Л –Є–Ј –Ї–ї–µ—В–Њ–Ї –Ї–Њ—Б—В–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –Є–ї–Є –Ї—Г–ї—М—В—Г—А—Л —В–Ї–∞–љ–µ–є, –њ—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–µ –њ–Њ –Њ–±—Л—З–љ—Л–Љ –Љ–µ—В–Њ–і–Є–Ї–∞–Љ —Б –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ–Љ —Ж–Є—В—А–∞—В–љ–Њ–є –≥–Є–њ–Њ—В–Њ–љ–Є–Є –Є —В–µ—Е–љ–Є–Ї–Є –≤—Л–ґ–Є–≥–∞–љ–Є—П —Д–Є–Ї—Б–∞—В–Њ—А–∞, –љ–∞ 15-20 —Б–µ–Ї. –Њ–њ—Г—Б–Ї–∞–ї–Є –≤ 0,25%-–љ—Л–є —А–∞—Б—В–≤–Њ—А —В—А–Є–њ—Б–Є–љ–∞, –љ–∞–≥—А–µ—В—Л–є –і–Њ 30¬∞–°. –Я–Њ—Б–ї–µ —Н—В–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В—Л —Б–њ–Њ–ї–∞—Б–Ї–Є–≤–∞–ї–Є –≤ 2√ЧSSC-–±—Г—Д–µ—А–µ (–≤ 1 –ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л —А–∞—Б—В–≤–Њ—А–Є—В—М 17,53 –≥ —Е–ї–Њ—А–Є—Б—В–Њ–≥–Њ –љ–∞—В—А–Є—П –Є 8,82 –≥ —Ж–Є—В—А–∞—В–∞ –љ–∞—В—А–Є—П), –∞ –Ј–∞—В–µ–Љ –њ–Њ–Љ–µ—Й–∞–ї–Є –≤ —В–µ—А–Љ–Њ—Б—В–∞—В –≤ —Б–≤–µ–ґ–µ–Љ —А–∞—Б—В–≤–Њ—А–µ —Н—В–Њ–≥–Њ –±—Г—Д–µ—А–∞, –≥–і–µ –≤—Л–і–µ—А–ґ–Є–≤–∞–ї–Є 1 —З–∞—Б –њ—А–Є 62¬∞ –°. –Я–Њ—Б–ї–µ —Н—В–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В—Л –Њ–Ї—А–∞—И–Є–≤–∞–ї–Є—Б—М –Ї—А–∞—Б–Є—В–µ–ї–µ–Љ –У–Є–Љ–Ј–∞ (1 –Љ–ї –Њ—Б–љ–Њ–≤–љ–Њ–≥–Њ –Ї—А–∞—Б–Є—В–µ–ї—П –У–Є–Љ–Ј–∞ –љ–∞ 50 –Љ–ї –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і—Л –њ—А–Є —А–Э 6,8-7,0) 5-10 –Љ–Є–љ., –Њ–њ–Њ–ї–∞—Б–Ї–Є–≤–∞–ї–Є—Б—М –≤ –і–Є—Б—В–Є–ї–ї–Є—А–Њ–≤–∞–љ–љ–Њ–є –≤–Њ–і–µ –Є –≤—Л—Б—Г—И–Є–≤–∞–ї–Є—Б—М. –Я—А–Є–≥–Њ—В–Њ–≤–ї–µ–љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л –њ–µ—А–µ–і –Њ–Ї—А–∞—Б–Ї–Њ–є –ґ–µ–ї–∞—В–µ–ї—М–љ–Њ —Е—А–∞–љ–Є—В—М –љ–µ –±–Њ–ї—М—И–µ –Њ–і–љ–Њ–≥–Њ –Љ–µ—Б—П—Ж–∞, –≤ –њ—А–Њ—В–Є–≤–љ–Њ–Љ —Б–ї—Г—З–∞–µ –Ї–∞—З–µ—Б—В–≤–Њ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є–∞–ї—М–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є —Г—Е—Г–і—И–∞–µ—В—Б—П.
|